Факты о фибробластах

Тело человека состоит из триллионов разнообразных клеток. Наиболее важными клетками самого большого органа в теле человека – кожи, являются фибробласты. Их называют клетками молодости, так как именно активная работа фибробластов способствует поддержанию молодости и красоты кожи.
Фибробласты
Зародышевые клетки соединительной ткани организма. Они участвуют в процессах регенерации и синтеза белков, наиболее важных для омоложения клеток дермы.
В организме человека фибробласты могут находиться в двух формах: активные и неактивные. Активный фибробласт имеет большой размер, отростки, овальное ядро и много рибосом. Такая клетка может делиться и интенсивно вырабатывать коллаген. Неактивные фибробласты называются также фиброцитами. Они являются высокодифференцированными клетками, которые образовываются их фибробластов, не имеют способности к делению, но принимают активное участие в синтезе волокнистых структур и заживлении ран. Неактивные фибробласты имеют несколько меньший размер, чем активные, и отличаются веретенообразной формой.
Все активные фибробласты разделяются на несколько структурно-функциональных типов, каждый из которых выполняет определенные функции:
— малодифференцированные фибробласты обладают выраженными пролиферативными свойствами, то есть, они активно размножаются и растут;
— юные фибробласты – более дифференцированные клетки, которые также способны к пролиферации, но в отличие от малодифференцированных, могут синтезировать коллаген и кислые гликозаминогликаны;
— зрелые фибробласты образуются из юных форм, практически не могут размножаться, и разделяются на три подтипа:
Фибробласты располагаются в среднем слое кожи человека – в дерме. Там они вырабатывают внеклеточный матрикс, компоненты которого и формируют своеобразный каркас кожи. Основными компонентами внеклеточного матрикса являются гликопротеины, протеогликаны и гиалуроновая кислота. Широко известный коллаген является превалирующим гликопротеином внеклеточного матрикса. Кроме того, фибробласты продуцируют также белки фибрин, эластин, тинасцин, нидоген и ламинин, которые используются в качестве «строительного материала» для кожи.
Еще один продукт синтеза фибробластов – это факторы клеточного роста, к которым относятся:
Основные функции фибробластов:
С возрастом в организме человека способности фибробластов в плане активного синтеза и пролиферации в тканях кожи снижаются, в результате чего происходит уменьшение содержания их главных компонентов — гиалуроновой кислоты, коллагена, эластина, сосудистой сети. Это отражается на внешнем виде кожного покрова.
Сегодня, благодаря успехам биотехнологии, появилась возможность естественным путем повлиять непосредственно на причину возрастного увядания кожных тканей. Этого удалось достигнуть способом обогащения ее собственными молодыми фибробластами, которые являются строителями внеклеточного матрикса.
Трансплантация в кожу лица собственных молодых клеток фибробластов способна эффективно и достаточно быстро активизировать процессы обновления и восстановления ее структуры. Аутологичные (свои) клетки не воспринимаются собственной иммунной системой как антиген (чужеродные) и, следовательно, организмом не отторгаются, а полноценно функционируют. Преимуществом клеточного омоложения является и то, что трансплантированные фибробласты долгое время (от полугода до полутора лет) сохраняют функциональную активность в части усиленного синтеза гиалуроновой кислоты, коллагена, эластина и других компонентов матриксной системы кожи. В течение этого срока постоянно продолжается улучшение ее состояния.
Такая методика аутотрансплантации фибробластов в косметологии получила официальное разрешение Росздравнадзора.
Фиброциты и фибробласты что это
В этой статье поговорим о функции дермальных фибробластов в коже, процессе их старения, а также о методе обновления и стимуляции регенерации кожи путём введения аутологичных фибробластов.
Введение
Роль дермальных фибробластов (ДФ) в обновлении кожи трудно переоценить, поскольку они являются основным клеточным компонентом соединительнотканной основы кожи, обеспечивающим её гомеостаз и морфофункциональную организацию. Очевидна актуальность этой темы для регенеративной (репаративной) косметологии, задача которой — омоложение кожи путём активации естественных процессов её восстановления.
Функции дермальных фибробластов
Из всех функций ДФ наибольшее внимание специалистов в области эстетической медицины и дерматологии сосредоточено на продуцировании этими клетками компонентов внеклеточного матрикса (ВКМ) дермы: коллагена, эластина, протеогликанов, особенно гиалуроновой кислоты (ГК), и структурных гликопротеинов.
Повышенный интерес именно к этой функции ДФ понятен: названные компоненты ВКМ формируют и поддерживают структуру кожи, обеспечивая её упругость и прочность. Причём ДФ не только синтезируют эти вещества, но и обеспечивают их катаболизм путём прямого фагоцитоза «отработанных» фрагментов фибрилл коллагена и эластина, секреции коллагеназы, гиалуронидазы и прочих ферментов.
Резидентные клетки ткани — образующиеся и постоянно пребывающие в этой ткани.
Другая, и не менее значимая для кожи, активность ДФ оставалась до недавнего времени несколько на обочине поля зрения врачей-практиков, однако сегодня она тоже становится объектом пристального внимания клиницистов. Речь идёт о целом ряде важнейших функций фибробластов. Так, продуцируя коллаген IV типа и ламинин, они влияют на формирование базальной мембраны. Фибробласты формируют строму, которая не только служит каркасом (опорой) для эпителия, но и регулирует структурную организацию и функционирование эпителиальных клеток. Они вырабатывают и выделяют факторы клеточного роста (KGF-1 — фактор роста кератиноцитов, GM–CSF — гранулоцитарно-макрафагальный колониестимулирующий фактор роста), иммунорегуляторные интерлейкины IL-6, IL-8. За счёт секреции факторов роста и интерлейкинов и непосредственного взаимодействия с эпителиальными клетками ДФ играют ключевую роль в регуляции эпидермального морфогенеза. Их паракринная активность служит основой для поддержания гомеостаза стволовых кератиноцитов.
Дермальные фибробласты секретируют факторы, влияющие на дифференцировку лимфоцитов, и факторы, регулирующие численность, миграцию и функции гранулоцитов и макрофагов, обеспечивая тем самым поддержание иммунитета кожи. Вырабатывая множество проангиогенных факторов, которые индуцируют дифференцировку и миграцию эндотелиальных клеток, ДФ способствуют образованию и стабилизации сосудов.
Они принимают участие в процессах нейроэндокринной регуляции кожи. Синтезируют биологически активные пептиды: гормоны, биогенные амины, нейропептиды и нейротрансмиттеры, идентичные таковым в центральной нервной и эндокринной системах, экспрессируют рецепторы андрогенов и эстрогенов, посредством которых осуществляется влияние этих гормонов на кожу человека.
Фибробласты, взаимодействуя с различными резидентными клетками: эпидермальными, эндотелиальными, нервными, жировыми, гемопоэтическими, участвуют практически во всех кожных процессах, в том числе — физиопатологических. Таким образом, ДФ представляют собой центральное звено в биологии кожи: они поддерживают не только гомеостаз ВКМ дермы, обеспечивая её ремоделирование и обновление, но и физиологическое состояние других слоёв и структур кожи.
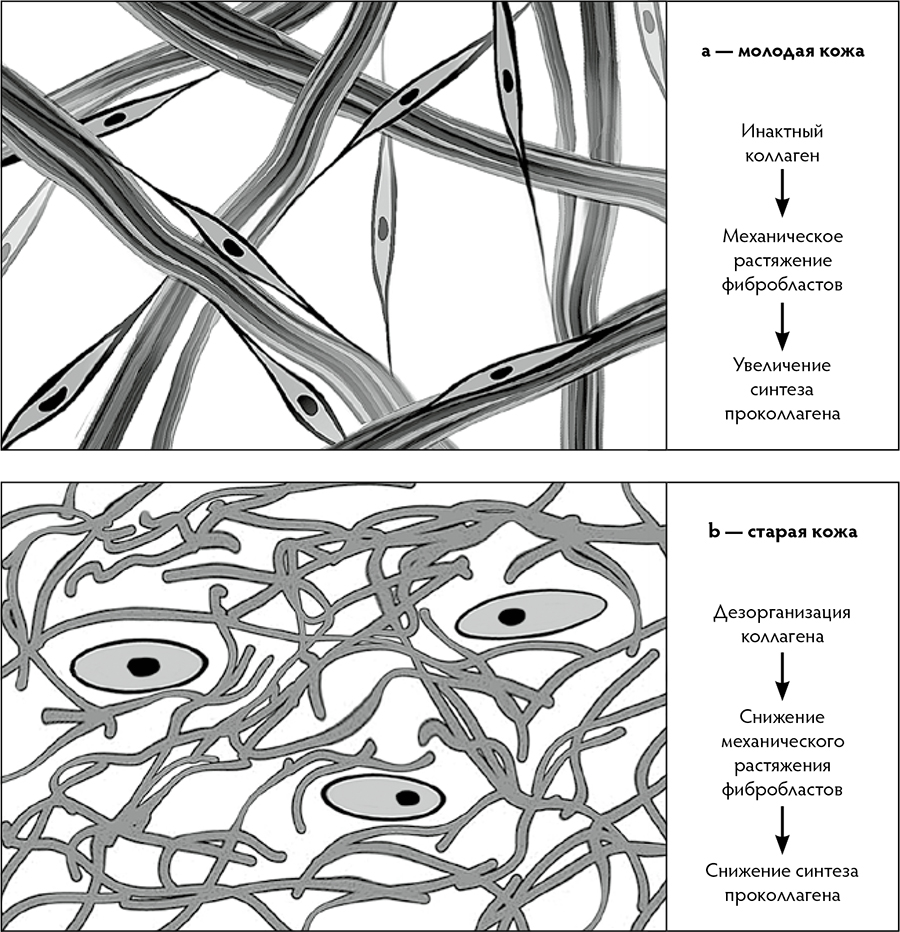 Рис. 1. Взаиморасположение фибробластов и коллагеновых волокон: a — в коже молодых людей (18–29 лет); b — в коже людей старшего возраста (старше 80 лет) (Varani J., et al. 2006, с изм.)
Рис. 1. Взаиморасположение фибробластов и коллагеновых волокон: a — в коже молодых людей (18–29 лет); b — в коже людей старшего возраста (старше 80 лет) (Varani J., et al. 2006, с изм.)
Инволюция дермальных фибробластов
По мере старения организма растёт число ДФ, в которых происходят возрастзависимые изменения, связанные преимущественно с непрерывным накоплением повреждений. Такие ДФ обозначают как «стареющие», или сенесцентные фибробласты (ДФст). Снижается их способность к ремоделированию и организации ВКМ, так как уменьшается синтез и секреция ими основных компонентов ВКМ. Нарушается эпидермальный гомеостаз из‑за негативных изменений в паракринных механизмах, связывающих эпидермис и папиллярный слой дермы. В популяции ДФст происходит увеличение секреции богатого цистеином ангиогенного индуктора — белка 61 (CYR61, или CCN1), стимулирующего продукцию провоспалительных цитокинов и матриксных металлопротеиназ (MMPs). CCN1 и MMPs поддерживают старение клетки путём негативной регуляции гомеостаза коллагена и увеличения его деградации.
Постепенно баланс между синтезом и деградацией коллагена нарушается в пользу последней. Повреждённый коллагеновый каркас ВКМ уже не может удерживать фибробласты в растянутом состоянии, свойственном молодым клеткам. Происходит своего рода коллапс фибробластов, что приводит к нарушению их функций. В процессе инволюции уменьшается численность фибробластов. Снижается и биосинтетическая активность этих клеток, нарушается баланс между синтезом и деградацией ВКМ дермы. В среднем, у пожилых / старых людей, по сравнению с молодыми, общее количество ДФ меньше на 35 %, продукция коллагена в коже снижена на 75 %, причём содержание коллагена уменьшается приблизительно на 1 % в год.
Все сенесцентные изменения в популяции ДФ приводят к постепенному и значительному снижению способности кожи к регенерации и обновлению. Как при этом меняется статус кожи? Уменьшается её толщина, снижается упругость и эластичность, появляются морщины, заломы, птоз тканей — кожа стареет.
Очевидно, что именно ДФ должны быть основной «мишенью» омолаживающих кожу косметологических методов. Так оно и есть: все современные терапевтические воздействия в целях коррекции возрастных изменений кожи — инъекционные, включая PRP-терапию, аппаратные (лазерные, радиочастотные, ультразвуковые) — направлены прежде всего на стимуляцию функциональной активности ДФ, как пролиферативной, так и биосинтетической. Особое место в этом ряду занимает применение аутологичных (собственных) дермальных фибробластов, или SPRS-терапия.
SPRS-терапия как метод оздоровления и обновления кожи
SPRS-терапия (SPRS — от англ. Service for Personal Regeneration of Skin — персонифицированный комплекс лечебно-диагностических процедур для восстановления кожи) — метод регенеративной медицины (РУ ФС №2009 / 308 от 21.07.2010). Его применение позволяет восполнить уменьшившуюся с возрастом популяцию ДФ за счёт введенных в кожу специализированных молодых и функционально активных клеток. Трансплантируются собственные фибробласты кожи пациента, поэтому они полноценно приживаются, интегрируют с резидентной клеточной популяцией и живут «по законам» дермы.
После трансплантации их биосинтетическая активность сохраняется не менее года, причём без какой‑либо дополнительной стимуляции. Как результат, увеличивается синтез коллагена и других компонентов ВКМ, благодаря чему происходит ремоделирование микроструктуры дермы, увеличение её толщины, уменьшение количества и глубины морщин.
Сенесценция — это состояние, при котором клетки с накопленными повреждениями ДНК и истощением механизмов её восстановления перестают делиться, теряют способность к ремоделированию и организации ВКМ.
Описание технологии
В специализированной лаборатории Института стволовых клеток человека, соответствующей международным стандартам GMP*, из 4 мм биоптата кожи пациента получают клеточный препарат, содержащий культивированные аутологичные ДФ (аутоДФ). На этапах клеточного процессинга происходит отбор и стимуляция только функционально активных ДФ, которые, несмотря на старение организма, сохраняют высокую способность к делению и синтезу важных для кожи компонентов. Дело в том, что пролиферативный потенциал всей популяции дермальных фибробластов взрослого человека в течение всей его жизни остаётся на довольно высоком уровне — благодаря наличию в фибробластическом диффероне стволовых / прогениторных клеток. Это клетки-предшественницы, которые отвечают за обновление клеточной популяции дермы. Как показали исследования in vitro, первичные культуры, полученные даже от очень пожилых людей (95 лет), содержат до 14 % митотически активных фибробластов.
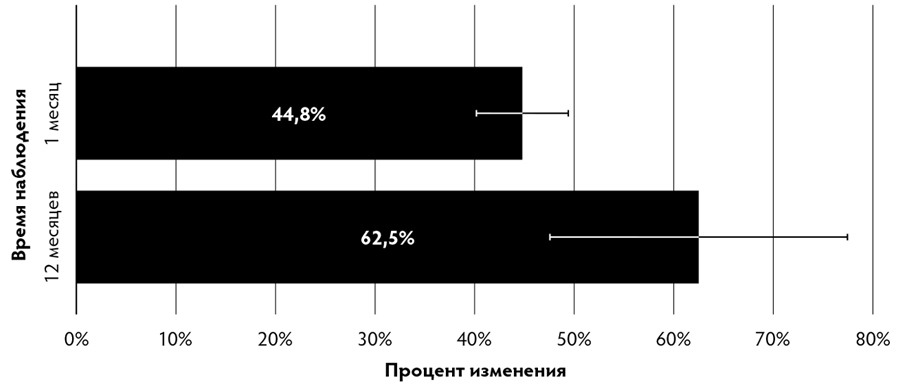 Рис. 3. Относительное изменение толщины кожи после применения аутоДФ (по результатам гистологических исследований).
Рис. 3. Относительное изменение толщины кожи после применения аутоДФ (по результатам гистологических исследований).
Данный факт подтверждается и нашими экспериментами: культуры фибробластов дермы, полученные от пациентов 18–82 лет, характеризуются довольно высокой эффективностью колониеобразования (45.0±9.5 %), независимо от возраста донора. Это даёт возможность из небольшого биоптата кожи любого взрослого человека получить необходимое для проведения терапии количество функционально активных клеток.
После трансплантации культивированных ауто ДФ в дерму их биосинтетическая активность сохраняется. В своей работе мы вводили полученный клеточный материал одновременно и в кожу лица, и в кожу за ушной раковиной, откуда затем проводили забор биоптата для гистологического изучения. Наши исследования показали:
Через 24 месяца после применения аутоДФ в дерме также регистрировали отдельные группы фибробластоподобных клеток [Рис. 4], продуцирующих коллагеновые волокна. Однако признаков созревания (утолщения) коллагена не наблюдали, из чего следует, что через два года после трансплантации аутоДФ интенсивность их синтетической активности (по сравнению с таковой в течение первого года после трансплантации) была снижена.
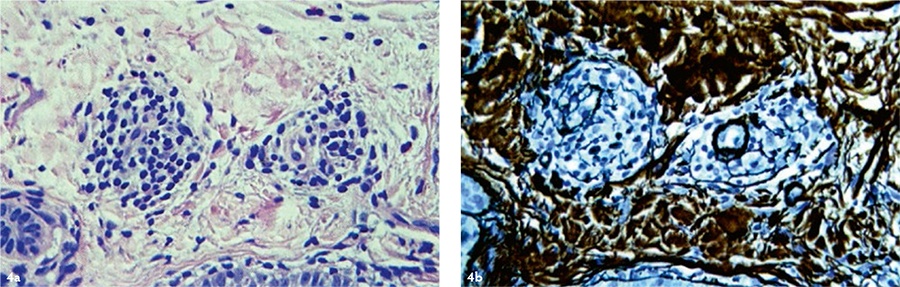 Рис. 4. Кожа пациента М. (54 г.) после применения аутоДФ через 24 месяца. Левая колонка — окраска гематоксилином и эозином, правая колонка — импрегнация нитратом серебра. Ув. ×200. Препараты и микрофотографии Р. В. Деева.
Рис. 4. Кожа пациента М. (54 г.) после применения аутоДФ через 24 месяца. Левая колонка — окраска гематоксилином и эозином, правая колонка — импрегнация нитратом серебра. Ув. ×200. Препараты и микрофотографии Р. В. Деева.
Скорее всего, это связано с уровнем физиологических потребностей дермы (известно, что для обеспечения её физиологического состояния достаточно незначительной функциональной активности фибробластов). По всей видимости, трансплантированные аутоДФ полноценно интегрировались с дермой, стали естественной составляющей её основной клеточной популяции — фибробластического дифферона — и находятся под контролем микроокружения.
Выводы гистологического исследования:
Выявленная на гистологическом уровне положительная динамика изменений кожи после применения аутоДФ полностью соответствует клинической картине. Уже через 10–14 дней после окончания курса SPRS-терапии отмечается повышение упругости кожи, уменьшение её рельефности (уменьшение выраженности морщин), улучшение цвета и контуров лица. Эффект имеет нарастающий характер. Так, если через месяц после инъекции на «хорошо» и «отлично» клинический результат оценили 88 % пациентов, то уже через три месяца и позже — 100 %. Врачи-исследователи через месяц определили результат как «хороший» и «отличный» у 86 % пациентов, через три месяца и позже — у 100 % пациентов. Положительные, прогрессирующие со временем (на протяжении как минимум 12 месяцев) изменения состояния кожи подтверждены нами и с помощью инструментальных методов исследования.
Заключение
Инновационная технология — SPRS-терапия, основанная на принципах регенеративной медицины, — позволяет восстанавливать утраченные с возрастом структуру и функции дермы за счёт уникального биологического механизма собственных коллагенобразующих клеток кожи пациента.
SPRS-терапия используется в отечественной эстетической медицине более восьми лет, зафиксировано более 1000 клинических наблюдений пациентов, из которых более 80 % повторно (два и более раз), пролечив одну область кожи, обращались в клиники для лечения этим методом кожи других областей.
Для врачебной практики важно, что SPRS-терапия является не только самостоятельным эффективным методом ремоделирования дермы, но и хорошей базовой основой для применения других косметологических вмешательств, направленных на стимуляцию синтетической активности фибробластов, что позволяет в течение длительного времени поддерживать кожу пациента в хорошем состоянии.
Авторы:
Алла Зорина, к. м. н., врач-биохимик, Москва.
Вадим Зорин, к. б. н., врач-биофизик, Москва.
Фиброциты и фибробласты что это
Фибробласты — ведущие клетки рыхлой соединительной ткани, продуцирующие компоненты межклеточного вещества. Это отростчатые, веретенообразные или распластанные клетки размером около 20 мкм. В них хорошо развиты органеллы внутренней метаболической среды. Ядро фибробласта овальной формы, содержит равномерно распыленный хроматин и 2-3 ядрышка. Цитоплазма отчетливо подразделяется на интенсивно окрашенную эндоплазму и слабо окрашенную эктоплазму. Цитоплазма фибробластов (особенно молодых) базофильна. В ней выявляется хорошо развитая эндоплазматическая сеть с большим количеством рибосом, прикрепленных к мембранам в виде цепочек по 10-30 гранул. Такая ультраструктура гранулярной эндоплазматической сети характерна для клеток, активно синтезирующих белок «на экспорт». Имеются также многочисленные свободные рибосомы, хорошо развитый комплекс Гольджи. Митохондрии — крупные, количество их невелико. Цитохимическими методами показано наличие в цитоплазме фибробластов ферментов гликолиза и гидролитических ферментов лизосом (особенно — коллагеназы). Менее активны окислительные ферменты митохондрии.
Опорно-двигательная система клетки обеспечивает их подвижность, изменение формы, прикрепление к субстрату, механическое натяжение пленки, к которой клетка прикрепляется в культуре. На клеточной поверхности имеется много микроворсинок и пузырчатых выростов. Фибробласты во взвешенном состоянии в жидкой среде имеют шаровидную форму. Распластанным фибробласт становится после прилипания к твердой поверхности, по которой он передвигается за счет псевдоподий.
Основная функция фибробластов — синтез и секреция белков и гликозаминогликанов, идущих на формирование компонентов межклеточного вещества соединительной ткани, а также выработка и секреция колониестимулирующих факторов (грану-лоцитов, макрофагов). Фибробласты долгое время сохраняют способность к пролиферации. Фибробласты, закончившие цикл развития, называются фиброцитами. Это долгоживущие клетки. Цитоплазма клеток обедняется органеллами, клетка уплощается, пролиферативный потенциал падает. Однако клетка не теряет способность участвовать в регуляции обменных процессов в ткани.
Межклеточное вещество. Состоит из фибриллярного и основного (аморфного) компонентов. Методами гистоавторадиографии с введением меченых аминокислот (3Н-пролин, 3Н-глицин и др.) установлено, что в полисомах фибробластов происходит синтез молекул белка. Фибробласты одновременно могут синтезировать несколько типов специфических белков и гликозаминогликаны. Для синтеза белка коллагена имеет существенное значение наличие витамина С, при недостатке которого коллагеногенез резко тормозится. Интенсивнее идет синтез межклеточного вещества в условиях пониженной концентрации кислорода. Одновременно с синтезом коллагена фибробласт разрушает примерно 2/3 этого белка с помощью фермента коллагеназы, что препятствует преждевременному склерозированию ткани.
Синтезированные молекулы проколлагена выводятся на поверхность фибробластов путем экзоцитоза. При этом осуществляется переход белка из растворимой формы в нерастворимую — тропоколлаген. Объединение молекул тропоколлагена в надмолекулярные структуры — коллагеновые фибриллы — происходит в непосредственной близости от клеточной поверхности благодаря действию особых веществ, выделяемых клеткой. В частности, на поверхности фибробластов обнаружен белок — фибронектин, выполняющий адгезивную и другие функции. Последующие этапы фибриллогенеза происходят путем полимеризации и агрегации тропоколлагена на ранее образованных фибриллах. При этом созревание коллагеновых волокон может протекать и без прямой связи с фибробластами.
Гликозаминогликаны являются регуляторами коллагенообразования и входят в состав основного (аморфного) компонента межклеточного вещества.
Фибриллярный компонент межклеточного вещества рыхлой соединительной ткани включает три типа волокон — коллагеновые, эластические и ретикулярные. Они имеют сходный механизм образования, однако отличаются друг от друга по химическому составу, ультраструктуре и физическим свойствам. Белок коллаген идентифицируется по аминокислотному составу и последовательности расположения аминокислот в молекуле коллагена. В зависимости от вариации аминокислот в полипептидной цепи, иммунных свойств, молекулярной массы и др. различают 14 и более разновидностей коллагеновых белков, которые входят в состав соединительной ткани органов. Все они составляют 4 основных типа, или класса, коллагена.
Коллаген 1-го типа встречается в соединительной и костной тканях, а также в склере и роговице глаза; II-го типа — в хрящевых тканях; III-го типа — в стенке кровеносных сосудов, в соединительной ткани кожи плода; IV-ro типа — в базальных мембранах.
Биологическая роль фибробластов в развитии опухолей

Аннотация
Среди всех клеток человеческого организма только фибробласты можно сравнить с тараканами по их способности к выживанию. Они в состоянии пережить тяжелый стресс, который обычно летален для других клеток, и только они могут быть культивированы из ткани после смерти организма или в процессе умирания ткани. Их устойчивость может быть обусловлена внутренними программами выживания и их пластичностью. Развитие злокачественной опухоли связано с фибробластами на всех стадиях, включая метастазирование. Они являются ключевым компонентом ответа организма на повреждение, вызванными опухолевыми клетками. Фибробласты, ассоциированные с опухолью (ФАО), становятся синтетическими машинами, которые продуцируют различные компоненты опухолей. ФАО играют роль в создании внеклеточного матрикса (ВКМ) и перепрограммировании метаболизма и иммунитета в микроокружении опухоли и ее адаптации к химиотерапии. Плейотропное действие ФАО на опухолевые клетки обусловлено их неоднородностью и пластичностью с разносторонним влиянием, зависящим от факторов среды.
Развитие злокачественных клеток запускает патологические процессы в организме, приводящие к формированию опухолевой ткани, которая содержит компоненты нормальных органов 4. Эти процессы, определяющие сложные гетерогенные взаимоотношения злокачественных клеток с нормальными клетками организма, известны как десмопластическая реакция, опухолевая строма или опухолевое микроокружение (ОМО). Опухолевая строма состоит из клеток иммунной системы, капилляров, базальной мембраны, активированных фибробластов и внеклеточного матрикса (ВКМ), окружающего злокачественные клетки [2]. На сегодняшний день стало совершенно очевидно, что развитие злокачественной опухоли и метастазирование опосредовано эффектами ОМО и зависит не только от автономных нарушений клетки [4, 8, 9]. Клетки иммунной системы, ангиогенез, напряжение кислорода, интерстициальное давление, ремоделирование ВКМ и метаболитов опухолевых клеток из ОМО заслужили особого внимания как важные детерминанты поведения клеток опухоли и прогрессии заболевания [1, 5, 10]. Основной компонент опухолевой стромы – фибробласты. Многие исследования за последнее время были направлены на изучение роли фибробластов в процессе развития злокачественной опухоли и метастазировании [11, 12]. Фибробласты, ассоциированные с опухолью были названы опухоль-ассоциированными фибробластами (ФАО), активированными фибробластами или активированными миофибробластами. Они могут включать также и мезенхимальные стволовые клетки (МСК). За прошедшее десятилетие ФАО прочно закрепили за собой роль ключевых компонентов в развитии злокачественной опухоли и новые исследования говорят об их участии во многих стромальных программах различных опухолевых процессов [13, 14]. Фибробласты обычно неподвижны и становятся активными только в ответ на повреждение ткани. Точный механизм их активации до сих пор остается невыясненным. Однако считается, что есть два типа активации фибробластов: “обратимый” и “необратимый”, частично определяющийся эпигенетическими факторами [15, 16] (РИС. 1). Активированные фибробласты были впервые описаны в механизме заживления ран и выявлены главным образом благодаря экспрессии α-гладкомышечного актина (α-ГMA; также известный как ACTA2) – белка цитоскелета, ассоциированного с клетками гладких мышц [17]. Из-за экспресии α-ГMA фибробласты называют миофибробластами 17. Активированные фибробласты являются также главным компонентом рубцов и важнейшим участником ответа на хронический процесс заживления ран, также известный как тканевой или органный фиброз [15, 22-25]. Постоянно продолжающийся (хронический) процесс заживления ран наблюдается при фиброзе органов и опухолевом росте [26], и он сильно отличается от соответствующего острого процесса [27, 28]. Несомненно, фибробласты являются самыми изменчивыми и наиболее изучаемыми клетками in vitro благодаря простоте их изолирования и культивирования. Они способны выживать при сильном стрессе и могут быть культивированы из тканей человека после его смерти [29]. В условиях беспощадного, высокодинамичного и опасного тканевого микроокружения, включая повреждение, вызванное химиотерапией или лучевой терапией, ФАО могут представлять постоянный вид стромальных клеток, который участвует в рецидиве опухоли. Несмотря на то, что они являются наиболее изучаемыми клетками в биологии, фибробласты остаются самыми мистическими и загадочными клетками. В последние годы всё больше информации появляется об их разнообразии и двойственной функции в развитии злокачественной опухоли.
Несмотря на схожесть, обусловленную очевидными идентичными чертами на клеточном уровне, нам всё ещё предстоит выяснить функциональные отличия между ФАО и фибробластами, ассоциированными с фиброзом (ФАФ) на молекулярном уровне. Более того, наши знания о пластичности фибробластов продолжают расти, формируя понимание того, какую роль она играет в функциональной вариабельности и сложности строения ОМО. В этом обзоре происхождение и функции фибробластов в физиологическом и патологическом перерождении паренхимы взрослого человека собраны воедино с особым акцентом на их функциональной гетерогенности в здоровом состоянии и при болезни.
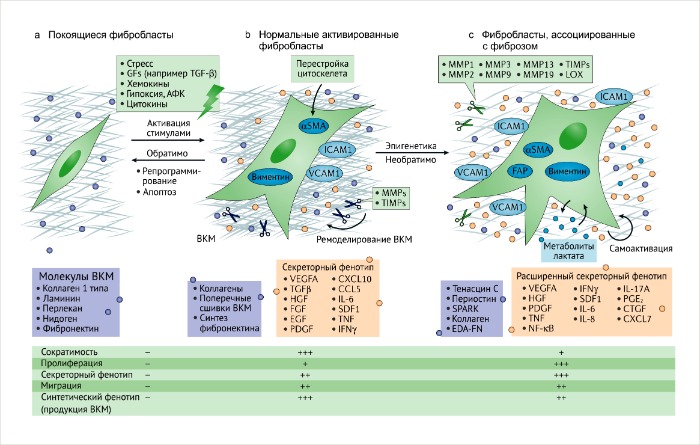
а. Покоящиеся или неактивные фибробласты – это инертные интерстициальные веретенообразные клетки, погруженные в физиологический внеклеточный матрикс (ВКМ).
b. В ответ на тканевое повреждение или ассоциированный с ним стимул, покоящиеся фибробласты обратимо активируются для обеспечения заживления и регенерации. Нормальные активированные фибробласты (НАФ) начинают экспрессировать актин гладких мышц (αSMA) и виментин, приобретая звездчатую форму.
Вместе с повышенной продукцией ВКМ, ремоделированием и перестройкой цитоскелета, фибробласты приобретают сократительные свойства. Полученные функции синтеза связаны с секрецией и миграцией, обуславливающими их активацию, вовлечение в клеточные процессы и пролиферацию. Обратимость такой активации может регулироваться репрограммированием или апоптозом активированных фибробластов, ассоциированных с процессом заживления, когда регенеративный процесс окончен.
с. Активированные фибробласты могут приобретать секреторный фенотип, специализируясь на ремоделировании ВКМ, аутокринной активации и динамическом иммуномодуляторном сигналинге. Этот процесс связан с постоянными и неутихающими повреждающими стимулами, такими как появление и развитие злокачественных очагов. Эпигенетическая регуляция может ограничивать регрессию таких активированных состояний. Фибробласты, ассоциированные с фиброзом (ФАФ) и опухолью (ФАО), могут приобретать повышенную пролиферативную активность и различный фенотип, тем самым увеличивая гетерогенность среды фиброзного и опухолевого микроокружения.
Определение и происхождение фибробластов
Первоначально Вирхов [30] (ок. 1858) и чуть позже Дюваль [31] использовали классические анатомические приемы и микроскопию, чтобы описать клетки, находящиеся в соединительной ткани. Фибробласты были первыми обнаруженными клетками соединительной ткани, которые продуцировали коллаген. В нормальных тканях они обычно являются одиночными клетками, которые находятся в интерстиции и иногда рядом с капиллярами, без каких-либо связей с базальной мембраной, но внедренные в фибриллярные структуры межклеточного матрикса. Фибробласты являются неэпителиальными, неиммунными клетками с вероятным мезенхимальным происхождением и составляют значительную часть разнородных компонентов соединительной ткани [32] (БОКС 1).
Фибробласты представляют собой классические веретенообразные клетки с потенцией к планарной полярности (РИС. 1). Фибробласты в нормальной ткани обычно рассматривают как медленнорастущие клетки с незначительной метаболической и транскрипционной активностью. При этом они находятся в спящем, бездействующем режиме. Покоящиеся фибробласты могут иметь много общих черт с предшественниками МСК (БОКС 2). Способность покоящихся фибробластов к активации и превращению в синтезирующие клетки (миофибробластами) было впервые выявлено при исследовании заживления ран [33] и позже при остром и хроническом воспалении и фиброзе тканей [17, 24, 34]. Нет никаких прямых доказательств того, что неактивные тканевые фибробласты у взрослых людей являются синтетически активными, кроме тех, что находятся в интерстициальном ВКМ.
Активированные фибробласты являются гетерогенной клеточной популяцией с различной экспрессией, зависящей от ткани, из которой они изолированы [235]. Количество клеточных пассажей и неидентичные условия культивирования могут также объяснить такую разнородность. Экспрессия гомеобоксных генов (HOX-генов) в фибробластах ограничена при культивировании [235], что подтверждает тот факт, что гетерогенность фибробластов отражает различные направления местной дифференцировки. Тщательное изучение собранных фактов говорит о том, что эмбриональные и развивающиеся ткани не имеют фибробластов, а, вероятно, наполнены мезенхимальными клетками. Как только развитие ткани завершено, большинство активных мезенхимальных клеток подвергается апоптозу, но некоторые (в любой ткани) превращаются в бездействующие фибробласты. Такие клетки наблюдал Вирхов [30] и позже назвал их фибробластами. В таких тканях, как взрослая сердечная мышца наблюдается большое количество бездействующих фибробластов. Таким образом, на сегодняшний день все свойства, приписываемые фибробластам, связаны с активированным фенотипом, а именно с их подтипом – миофибробластами. Одна из возможных функций бездействующих фибробластов – это их способность дифференцироваться в активную форму и, таким образом, основываясь на соответствующем стимуле, давать начало росту мезенхимальным линиям, включая хондроциты, адипоциты и эндотелиоциты. В этом отношении бездействующие фибробласты могут быть рассмотрены как взрослая форма тканевых мезенхимальных стволовых клеток. (rMSCs). Очевидно, что такие стволовые клетки отличаются от мезенхимальных стволовых клеток костного мозга (классических BM-MCS). Каким образом два типа мезенхимных стволовых клеток различаются по своим функциям, еще предстоит узнать.
αSMA, α‑гладкомышечный актин; ECM, внеклеточный матрикс; iPSCs, индуцированные плюрипотентные стволовые клетки; PDGFRβ, тромбоцитарного фактора роста рецептор-β; RASAL1, активатор RAS-белка 1.
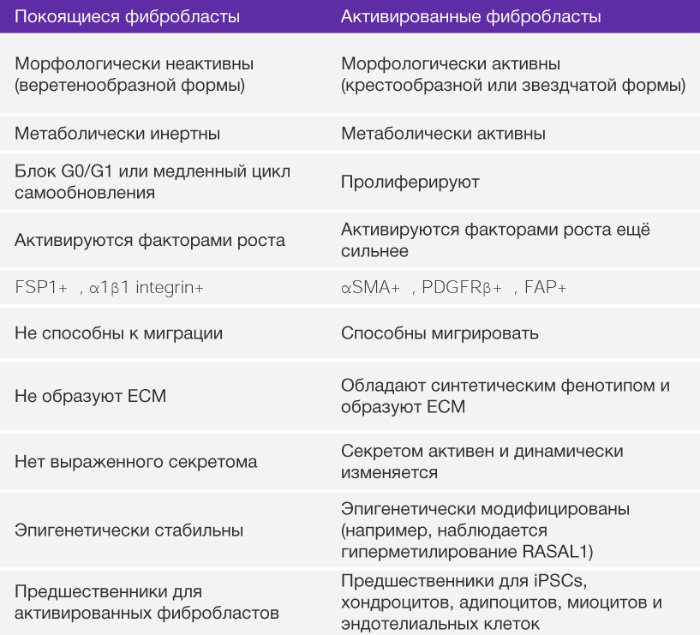
На самом деле, более точное определение фибробластов – покоящиеся мезенхимальные клетки, обладающие потенциальной возможностью активации соответствующим стимулом и превращением в МСК (БОКС 2). Такие покоящиеся мезенхимальные клетки являются довольно редкими клетками интерстиция, способные к пролиферации при стимуляции факторами роста, такими как трансформирующий фактор роста бета (TФР-β), тромбоцитарный фактор роста (ТрФР) и интерлейкин-6 (ИЛ-6), и другие [11, 12] (БОКС 3). Исходя из этого нового определения можно предположить, что большинство свойств, приписываемых фибробластам, на самом деле являются свойствами “активированных” фибробластов, миофибробластов и МСК. Фибробласты, полученные из места заживления раны или из фибринозной ткани, секретируют повышенные количества элементов ВКМ и пролиферируют чаще, чем их товарищи, изолированные из здоровых органов [35, 36]. Такая повышенная активность связана с их активацией [35] (РИС. 1). После активации они начинают синтезировать ВКМ, цитокины и хемокины, мобилизуют иммунные клетки и прилагают физические силы для модификации архитектуры тканей [37, 38]. Хотя обнаружение фибробластов во многих тканях и не представляет сложности в наше время, их идентификация в эмбриональных тканях неоднозначна. В целом, покоящиеся фибробласты не встречаются в ходе эмбрионального развития и впервые отмечаются в дифференцированных тканях и органах. У взрослых людей остается неизвестным, вносят ли вклад в популяцию активированных фибробластов МСК или мезенхимальные предшественники моноцитов (фиброциты).
Некоторые факты говорят о том, что МСК из костного мозга (КМ-MСК) могут усиливать метастатическую активность клеток рака молочной желез [39]. Например, КМ-МСК составляют значительную часть ФАО с протуморогенными функциями при раке желудка, индуцированном воспалением [40]. Фиброциты, или клетки костномозгового происхождения CD45+ (CD45 также известен как PTPRC), продуцирующие коллаген I типа, могут способствовать образованию в рубце фиброзной ткани благодаря накоплению коллагена и продукции провоспалительного секретома [41]. Остается одна гипотеза – МСК могут быть просто бездействующими фибробластами, которые могут стать активированными, или МСК с мультилинейным потенциалом. Доподлинно известно, что активированные фибробласты при культивировании могут стать адипоцитами, эндотелиоцитами или хондроцит-подобными клетками и при индукции становятся индуцированными плюрипотентными стволовыми клетками (ИПСК) 43 (РИС. 2). Важно помнить, что бездействующие фибробласты не используются для таких экспериментов по индукции пластичности.
Заживление ран, фиброз и опухоль
Острый период заживления ран
Любое повреждение функциональной паренхимы приводит к какому-либо ответу организма. К повреждающим стимулам относят механическую травму, ущерб от радиации, экстремальной температуры, токсинов, патогенов и метаболических нарушений. Ответ организма начинается незамедлительно после нанесения повреждения функциональной паренхиме и заключается в регенерации поврежденных участков клетки и восстановлении тканевого гомеостаза [21, 34, 37, 46, 47]. Такое заживление ран запускается в ответ на различные типы острых повреждений. Классический процесс заживления включает воспаление, активацию ангиогенеза иммунными клетками и фибробластами и отложение ВКМ [12, 26, 34]. Многие составляющие ВКМ и базальных мембран – такие как коллагены I, III, IV и V типов, различные ламинины и фибронектины – продуцируются активированными фибробластами или миофибробластами [37, 48]. Миофибробласты также являются главным источником протеаз, которые отвечают за деградацию ВКМ, включая матричные металлопротеазы (ММП), что подчеркивает их ключевую роль в сохранении гомеостаза ВКМ благодаря контролю за его обновлением [49]. Миофибробласты были впервые выделены при заживлении кожной раны. Именно их активность была связана с процессом стягивания кожи [32, 37]. Миофибробласты, индуцированные ТФР-β–опосредованным сигнальным путем, пролиферируют и экспрессируют виментин и α-ГМА [17, 18]. Кроме этого, активированные фибробласты играют важную роль в сохранении гомеостаза прилегающего эпителия с помощью секреции факторов роста и прямых взаимодействий между эпителиальными и мезенхимальными клетками [50] (БОКС 1). После заживления количество активированных фибробластов значительно уменьшается благодаря апоптозу, и изначальный фенотип покоящихся фибробластов, вероятно, восстанавливается [37] (РИС. 1). Такая обратимость является важным признаком восстановления ткани, связанного с заживлением.
Тканевой фиброз
Если повреждающий фактор действует постоянно, как при воздействии хронических физических, токсических, метаболических и аутоиммунных факторов, регенеративный ответ идет безостановочно и приводит к хроническому процессу заживления, также известному как фиброз. Таким образом, тканевой фиброз можно рассматривать как повреждение, которое тормозит процесс заживления из-за постоянной активации механизмов регенерации. Частично это регулируется эпигенетическими механизмами в активированных фибробластах, что усиливает активность антиапоптотических сигнальных путей и пролиферацию клеток, приводящую к генерации гиперактивированных фибробластов. Патологическое ремоделирование и фиброз в тканях могут повышать активность ФАФ с различным происхождением, маркерами активации и функциями 54.
Опухолевый фиброз
Хронический процесс регенерации ткани также осуществляется при генетическом повреждении функциональной паренхимы при злокачественной опухоли. В этом отношении опухоли можно рассматривать как «язвы, которые никогда не заживают» (ССЫЛКА 26). Постоянное возникновение и наращивание массы опухолевых клеток отражает прогрессирующее повреждение ткани, которое вызывает хронический процесс заживления по отношению к опухолевым клеткам. Это приводит к хронической ответной реакции организма, известной как опухолевый фиброз или стромообразование. Термин “опухолевый фиброз” аналогичен термину “десмопластическая реакция”, “опухолевое стромообразование” или ОМО. Роль миофибробластов в заживлении раны хорошо изучена, но их функциональное значение в прогрессии злокачественной опухоли и метастазировании представляется сложным и неоднозначным, одновременно и стимулирующим злокачественную опухоль, и тормозящим [12, 55, 56]. В 1970-х были выполнены новаторские исследования, которые позволили установить, что злокачественные клетки могут мобилизовать активированные фибробласты, сходные с фибробластами, ассоциированными с процессом заживления ран 61. Вовлечение стромальных фибробластов в опухоль значительно регулируется факторами роста, синтезируемыми злокачественными клетками и инфильтрированными иммунными клетками. ТФР-β, ТрФР и фактор роста фибробластов типа 2 (ФРФ-2) являются ключевыми медиаторами в активации фибробластов при остром и хроническом процессах повреждения ткани [62]. Мобилизация активированных фибробластов при многих видах злокачественных опухолей зависит от ТФР-β [63, 64]. Локальная пролиферация и инвазия ФАО стимулируется ТФР-β, представленными в опухолевом микроокружении. Существует еще одно предположение, что активация фибробластов является ответом организма на противодействие злокачественному роста и попыткой элиминации опухоли [11, 56, 62, 65]. ТрФР, секретируемый злокачественными и стромальными клетками (включая фибробласты), может активировать и индуцировать пролиферацию фибробластов и коррелировать с прогрессией злокачественной опухоли [66]. Точный механизм разрастания опухолевой стромы и опухолевого фиброза остается неизученным. На ранних стадиях образования солидных опухолей, злокачественные клетки формируют неопластические очаги, которые возникают в микроокружении эпителия [10, 67]. Многие исследования показали, что карцинома in situ уже включает в себя “реактивный” опухолевый фиброз [2, 68]. Ведутся оживленные дебаты по поводу значения реактивной опухолевой стромы в прогрессии злокачественной опухоли, где спорным является вопрос о ее роли в защите от инвазивных и злокачественных изменений в карциноме in situ [69].
В целом, действие опухолевой стромы рассматривается как стимулирующее возникновение злокачественной опухоли и ее развитие. Точный клеточный и молекулярный механизмы, которые осуществляют превращение карциномы in situ в инвазивный рак, неизвестны, но большинство ученых сходится во мнении, что опухолевая строма, скорее всего, является индуктором этого процесса. В этом отношении миоэпителиальные клетки, которые имеют некоторые общие черты с миофибробластами протоков молочных желез, содержащих злокачественные клетки, могут играть регуляторную роль в прогрессии опухоли [70].
На ранних этапах неоплазии воспалительные сигналы, появившиеся, возможно, в результате патологического преобразования ткани, могут стимулировать провоспалительные и опухоль-стимулирующие функции фибробластов. Секреция ИЛ-1β иммунными клетками в ранних опухолевых очагах ведет к активации в фибробластах сигнального пути транскрипцио́нного фактора NF-κB, результатом которого является образование проопухолевого секретома [71].
Создание ран у предрасположенных к злокачественным новообразованиям куриц, инфицированных вирусом саркомы Рауса, приводит к появлению инвазивной карциномы в толще поврежденной ткани [72]. В генномодифицированных мышах с экспрессией jun-онкогена также отмечается связь между нанесением наружного повреждения и появлением злокачественной опухоли [73]. Хотя эта связь остается неизученной у млекопитающих, во многих эпидемиологических и клинических исследованиях выдвинута гипотеза, что тканевой фиброз в таких органах, как печень, легкие и поджелудочная железа, может повысить риск появления карцином 76. Развитие гепатоцеллюлярной карциномы безусловно связано с предшествующим фиброзом печени [77, 78]. Нормальная строма в большинстве органов содержит небольшое число покоящихся фибробластов, погруженных в физиологический ВКМ [2], в то время как реактивная опухолевая строма или фиброзная ткань обычно наполнены большим количеством активированных фибробластов, которые обычно экспрессируют α-ГМА или белок-активатор фибробластов (БАФ) и демонстрируют повышенное отложение коллагена, фибрина и других составляющих ВКМ по сравнению с бездействующими фибробластами [6, 11, 13]. Привлечение новых капилляров и аккумуляция иммунных клеток в ОМО регулируется многими факторами роста, такими как фактор роста эндотелия сосудов А (ФРЭС-A), ТрФР, эпидермальный фактор роста (ЭФР), ИЛ-6 и ИЛ-8, наряду с некоторыми другими. Вдобавок к продукции ФРЭС-A злокачественными клетками, ее также осуществляют активированные фибробласты [79]. ФРЭС-A, также известный благодаря своей способности увеличивать проницаемость сосудов, играет центральную роль в транссудации жидкости из сосудов, приводя к повышенной реактивности периваскулярной области, содержащей фибрин и тромбоциты. Они, в свою очередь, стимулируют аккумуляцию иммунных клеток, пролиферацию эндотелиоцитов и активированных фибробластов [80]. Опухолевая строма насыщается белками ВКМ, такими как коллаген I типа и фибронектин, которые инициируют ангиогенез 80. С ростом опухоли и приобретением инвазивности, стромальный компонент также увеличивается. Опухолевая строма на поздних стадиях злокачественной опухоли содержит повышенное количество различных типов коллагена, ламининов, фибронектинов, протеогликанов, периостинов и тенасцина-С, наряду с другими веществами [5, 83]. Периостин и тенасцин-С, частично продуцируемые активированными фибробластами, обычно отсутствуют в нормальных тканях молочной железы у взрослых, но они экспрессируются при раке груди [2, 84-87]. Многие недавние исследования продолжают указывать на то, что активированные фибробласты регулируют развитие злокачественной опухоли благодаря активному действию их секретома, который включает факторы роста и ВКМ.
Функция фибробластов и их гетерогенность
На сегодняшний день совершенно ясно, что многие различные маркеры позволяют идентифицировать активированные фибробласты. К ним относятся: специфический белок фибробластов (СБФ-1, также известный как S100A4), виментин, α-ГMA, БАФ, ТрФР-α, ТрФР-β, десминовые и дискоидиновые домен-содержащие рецепторы типа 2. Также важно заметить, что рецепторы ТрФР (ТрФР-α and ТрФР-β) и связанный с ними сигнальный путь могут служить мишенями для фармакологических средств 90. СБФ-1 является надежным маркером для определения покоящихся непролиферирующих (Ki67-негативных) фибробластов в интерстиции [91]. Важно отметить, что ни один из этих маркеров не является строго специфичным для покоящихся или активированных фибробластов. Например, СБФ-1 также идентифицирует макрофаги и, возможно, другие иммунные клетки; кроме того, экспрессируется некоторыми злокачественными клетками [92, 93]. БАФ также присутствует в подгруппе иммунных клеток CD45+ [94]. Десмин и ТрФР-β экспрессируются периваскулярными клетками [95]. Таким образом, при использовании этих маркеров содержание, морфология и пространственное распределение клеток должны приниматься во внимание для идентификации покоящихся и активированных фибробластов. Также вероятно, что многие активированные фибробласты не экспрессируют все эти общепризнанные маркеры в одно и то же время [96, 97], что повышает степень их неоднородности (РИС. 4). Остается неизвестным, выполняет ли какие-либо уникальные функции каждая из подгрупп активированных фибробластов, идентифицированных несколькими перекрывающимися или неперекрывающимися маркерами. Маркеры ФАО могут также быть связаны с клетками, обладающими разнообразными и, возможно, противоречащими функциями даже в одном специфическом опухолевом микроокружении. Потеря экспрессии кавеолина-1 при раке молочной железы может определять развитие метаболически измененных фибробластов с про-опухолевыми функциями [98, 99], в то время как высокая экспрессия кавеолина-1 у ФАО может обеспечить опухолевую инвазию посредством ремоделирования ВКМ [100]. Покоящиеся фибробласты, предположительно, также как и Т-лимфоциты, способны дифференцироваться в разнообразные подгруппы активированных фибробластов, которые, скорее всего, будут иметь различную активность (РИС. 5).
Чтобы идентифицировать такие подгруппы, следует использовать набор различных маркеров клеточной мембраны для детекции. Таким образом, необходимо проводить соответствующие исследования в целях определения их специфической активности. Гетерогенность фибробластов, в частности активированных, может также зависеть от происхождения предшественников фибробластов. Активированные фибробласты имеют костномозговое происхождение; их предшественниками в целом могут являться МСК, эндотелиоциты, звездчатые клетки печени и поджелудочной железы, покоящиеся тканевые фибробласты, и, возможно, некоторые эпителиальные клетки [4, 11, 22, 40]. В зависимости от их происхождения, функции таких активированных фибробластов могут быть различными и уникальными. Возможно, будущие исследования смогут ответить на эти интригующие вопросы.
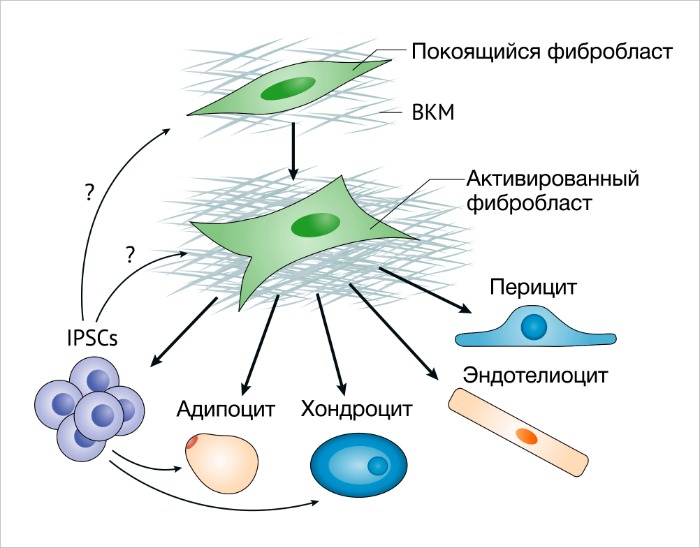
Фибробласты являются выоскопластичными клетками и обладают мультипотентностью. Активированные фибробласты быстро трансдифференцируются в хондроциты, миоциты и адипоциты, эндотелиоциты и другие клетки 42. Активированные фибробласты могут приобретать периваскулярные и васкулярные функции. Индуцированные плюрипотентные стволовые клетки были первыми клетками, полученными при помощи активированных фибробластов, которые до сих пор являются их самым насыщенным источником [42]. Пластичность фибробластов также может определять их гетерогенные функции.
ECM – внеклеточный матрикс.
Эпигенетическая регуляция фибробластов
В большинстве карцином активированные фибробласты считаются главными регуляторами многих различных стромальных программ и сигнальных путей в опухолевых клетках [20]. Сигналы, которые регулируют превращение покоящихся фибробластов в ФАО в опухолях, вероятно, сложны, но в культуре некоторые фенотипические черты, характерные для ФАО, могут быть индуцированы под действием ТФР-β [18]. Несмотря на это, новые научные данные говорят о том, что необратимая активация фибробластов может быть вызвана эпигенетическими альтерациями [15, 16, 101, 102]. При развитии опухоли — процессе, отличающемся от процесса заживления ран, но имеющем сходство с фиброзом, фибробласты остаются постоянно активными. В последние годы было сделано предположение, что при острых ситуациях эффект факторов роста на активацию фибробластов имеет обратимый характер. Но при хронизации тканевого фиброза и опухолевого стромообразования активированные фибробласты получают уникальные свойства, которые не наблюдаются у фибробластов, ассоциированных с заживлением ран или при остром повреждении (РИС. 1). Исследования опухолей молочной железы у человека показывают, что стромальные клетки (преимущественно фибробласты) имеют уникальные эпигенетические изменения, которые не наблюдаются в фибробластах нормальной ткани молочной железы [103].
Долгое время считалось, что ФАФ и ФАО могут быть легко культивированы из опухолевой ткани и что они пролиферируют намного быстрее, чем активированные фибробласты, полученные из нормальной ткани, или из ткани с острым повреждением, или из заживающей раны. При обработке ФАФ с помощью деметилируюшего агента 5-азацитидина их пролиферативная активность падала, синтез коллагена I типа и экспрессия α-ГMA снижались. Гиперметилирование RAS-подобных белков-активаторов (RASAL1), наряду с частичной инактивацией RASAL1, приводит к реактивации RAS в почечных ФАФ, но не в тех фибробластах, что изолируются из нормальной ткани или ассоциированы с острым повреждением почек [104]. Это исследование предоставляет первое доказательство роли эпигенетических альтераций в необратимой активации ФАФ. При фиброзе легкого эпигенетические модификации могут определить резистентность ФАФ к FAS (также известному как TNFRSF6)-опосредованному апоптозу [105] и дифференцировке миофибробластов [106, 107]. Стромальные клетки простаты с гиперэкспрессией эпигенетических регуляторов высокомобильной группы AТ-петли 2 (HMGA2) играли важную роль в индукции интраэпителиальной неоплазии [108]. Эпигенетический переключатель, ответственный за регуляцию ингибиторного фактора лейкемии (ИФЛ), может привести к активизации проинвазивных функций ФАО благодаря повышенной активности сигнального трансдуктора Янус-киназы 1 (JAK1) и активатора транскрипции 3 (STAT3). Двойное ингибирование ДНК-метилтрансферазы и активности Янус киназы поддерживает неинвазивный фенотип ФАО [109]. Будущие исследования, возможно, раскроют гораздо больше таких эпигенетических контрольных узлов в генерации ФАО.
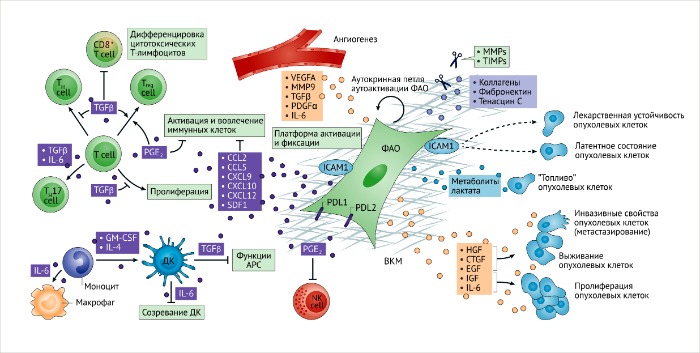
ФАО и их секретом участвуют в ремоделировании опухолевой стромы. Внеклеточный матрикс (ВКМ) вместе с клеточными компонентами опухолевого микроокружения сильно изменяются и репрограммируются фибробластами, ассоциированными с опухолью (ФАО). Секреторные функции регулируют иммунную перестройку (слева), самоактивацию (посередине) и связывают опухолевые клетки (справа), инициируя или ограничивая их рост, выживание или сопротивляемость к терапии. Метаболическая перестройка в ФАО может также поддерживать опухолевое микроокружение и стимулировать адаптацию опухолевых клеток к опухолевому росту. ФАО также могут сильно влиять на опухолевый ангиогенез, который косвенно влияет на вовлечение иммунных клеток и их активацию, миграцию опухолевых клеток и их инвазивные свойства. Внутриклеточная молекула адгезии (ICAM1), которая экспрессируется ФАО, может служить в качестве депо для активации и репрессии иммунных клеток и лиганда 1 белка программируемой клеточной смерти (PDL1), а экспрессия PDL2 на ФАО может влиять на иммуносупрессивные функции. Секретом ФАО имеет потенциальный ремоделирующий эффект на противоопухолевый иммунитет, затрагивая врожденные механизмы активации иммунных клеток, и поляризуя адаптивный иммунный ответ.
APC, антигенпрезентирующие клетки; CCL, хемокины с C-C мотивом; CTGF, фактор роста соединительной ткани; СXCL, хемокины с C-X-C мотивами; DC, дендритные клетки, EGF, эпидермальный фактор роста; GM-CSF, колониестимулирующий фактор роста гранулоцитов и макрофагов; HGF, гепатоцитарный фактор роста; IGF, инсулиноподобный фактор роста; IL, интерлейкин; MMP, матриксные металлопротеазы; NK, натуральные киллеры; PDGFα, фактор роста тромбоцитов; PGE2, простагландин E2; SDF1, фактор роста стромальных клеток; TGFβ, трансформирующий фактор роста; TH, Т-хелперы; TIMPs, тканевые ингибиторы металлопротеиназ; Treg cells, регуляторные Т-клетки; VEGFA, фактор роста эндотелия сосудов А.
Фибробласты, ассоциированные с опухолью: положительные регуляторы развития злокачественной опухоли
ФАО и канцерогенез
Активированные фибробласты, изолированные из опухолей организма человека, проявляют различные свойства по сравнению с фибробластами, полученными из нормальных органов [32] (РИС. 1). Важно помнить, что программы активации фибробластов могут быть индуцированы искусственно, с помощью изменений условии среды. Несмотря на это, существуют некоторые фундаментальные различия между культурированными нормальными фибробластами и аналогичными клетками, полученными из органов с тканевым фиброзом или из опухолевой ткани. ФАФ и ФАО пролиферируют гораздо быстрее, чем нормальные активированные фибробласты (НАФ). ФАО и ФАФ проявляют повышенную миграционную способность, обладают активным сигналингом фактор-индуцируемого аутокринного роста и высоким уровнем молекулярной секреции, что включает в себя выделение факторов роста и хемокинов (РИС. 1). Такое различие может быть обусловлено обнаруженными эпигенетическими изменениями, вероятно, как следствие влияния способствующей их активации внешней среды: в ФАФ и ФАО, но не в НАФ. Во многих экспериментах с совместным культивироваванием ФАО усиливали развитие опухоли, в отличие от НАФ [111]. Первоначально было продемонстрировано, что когда трансформированные симианским вирусом 40 (SV40) эпителиальные клетки простаты были смешаны с НАФ и ФАО, и эти смеси были привиты мышам, то именно ФАО (в отличие от НАФ) приводили к образованию неоплазий, похожих на интраэпителиальную опухоль [112]. ФАО также были способны индуцировать инвазию неинвазивных опухолевых клеток [113]. Подобные исследования были проведены и с другими видами опухолей [111]. Способность ФАО влиять на процесс развития опухоли зависит от их способности индуцировать ангиогенез, опосредованно через фактор стромальных клеток 1 (ФСК-1; также известен как CXCL12) или вовлечение эндотелиоцитов костномозгового происхождения [111]. Некоторые секретируемые ФАО молекулы-регуляторы имеют проонкогенный эффект (БОКС 3). Повышение экспрессии фактора теплового шока 1 (ФТШ-1) в ФАО может содействовать ФТШ-1-опосредованной программе в злокачественных клетках, поддерживая проопухолевое влияние ОМО [114]; а активация Yes-ассоциированного белка в ФАО усиливает уплотнение ВКМ и инвазивность злокачественных клеток [115]. Деактивация сигнальный путей, опосредованных Notch и p53 в ФАО, в дальнейшем усиливает пролиферацию этих клеток [116]. Для понимания того, как эти пути деактивируются в ФАО, необходимы дальнейшие исследования. Активированные фибробласты продуцируют протеазы, деградирующие ВКМ — такие как ММП 118. Подвижность и инвазивность злокачественных клеток регулируются ММП. Стромелизин-1 (также известный как MMП-3) синтезируется активированными фибробластами в больших количествах и расщепляет Е-кадгерин, стимулируя в прилегающих злокачественных клетках эпителиально-мезенхимальный переход (ЭМП) и инвазионную способность [120]. Кроме того, активированные фибробласты синтезируют MMП-1, который также индуцирует инвазивность [117]. Далее, делеция рецептора TФР-β типа II в фибробластах, экспрессирующих СБФ-1, приводит к развитию инвазивной плоскоклеточной карциномы кардиального отдела желудка или к интраэпителиальной неоплазии, зависимой от гепатоцитарного фактора роста (ГФР), продуцируемого фибробластами [8]. Продуцируемые фибробластами экзосомы также являются положительными регуляторами развития опухоли и стромального перерождения, за счет регуляции активности фибробластов и химиорезистентности 123. Фибробласты, у которых отсутствуют четыре компонента тканевого ингибитора семейства металлопротеинкиназ, образуют экзосомы с повышенной экспрессией MMП и активностью дизинтегрина и белка с доменом металлопротеиназной (ADAM10), связанные с большей подвижностью злокачественных клеток, метаболическим перепрограммированием и индукцией свойств опухолевых стволовых клеток [125].
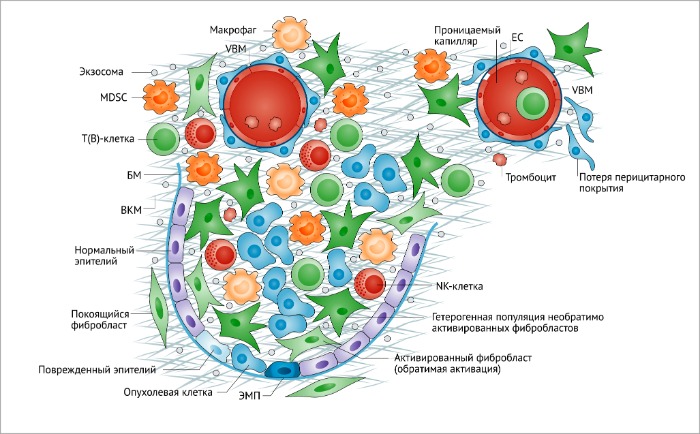
Фибробласты – гетерогенная популяция опухолевого микроокружения. Фибробласты, составляющие доминирующий клеточный компонент опухолевого микроокружения, которое, в свою очередь, состоит из инфильтрата иммунных клеток, нормального и поврежденного эпителия, неопластических эпителиальных клеток, перицитов и сосудистой базальной мембраны. Фибробласты, ассоциированные с опухолями (ФАО) являются гетерогенной популяцией необратимо активированных фибробластов с различными функциями.
BM, базальная мембрана; ЕС, эндотелиальные клетки; ЕМТ, эпителиально-мезенхимальный переход; MDSC, клетки-супрессоры миелодного происхождения; NK, натуральные киллеры.
Преобразование ВКМ фибробластами также может играть роль в создании и поддержании ниши опухолевых стволовых клеток. Фибробласты могут быть использованы опухолевыми стволовыми клетками для индукции среды по поддержанию своей популяции клеток. Экспрессия периостина (POSTN) фибробластами, вероятно, носит лимитирующий характер для стромального преобразования, мобилизации WNT-лигандов и поддержания ниши опухолевых стволовых клеток, регулируемой сигнальным путем WNT [126].
Повышение активности WNT-пути обнаружено во многих стволовых опухолевых клетках кишки, близко расположенных к ФАО, что, вероятно, вовлекает ГФР, синтезируемый такими фибробластами, в регуляцию создания ниши опухолевых стволовых клеток [127]. В случае раке легкого опухолевые стволовые клетки и клетки-инициаторы злокачественного роста являются сильными активаторами фибробластов благодаря экспрессии тромбоспондина-2, усиливающего процесс метастазирования [128]. Паракринная сигнализация между опухолевыми клетками и фибробластами задействует инсулиноподобный фактор роста II (ИФР-2), который также является производным фибробластов, и сигнальный путь, связанный с рецептором ИФР-1 в стволовых опухолевых клетках, усиливающий экспрессию Nanog и формирующий стволово-подобный фенотип опухолевых клеток [129].
Роль ФАО в метастазировании
Активированные фибробласты являются важными медиаторами вторичного роста опухоли в процессе метастазирования. В сайте первичной локализации опухоли ФАО способны усиливать интенсивность метастазирования благодаря высвобождению факторов роста и цитокинов в кровоток для стимуляции (косвенной или прямой) роста и инвазивных способностей злокачественных клеток в отдаленные структуры [114, 130, 131]. ФАО могут также влиять на уплотнение ВКМ в первичных опухолях, увеличивая инвазивность злокачественных клеток; кроме этого, они способны участвовать в образовании каналов ВКМ с целью перемещения по ним неопластически измененных клеток [133]. Фибробласты, ассоциированные с метастазированием (ФАМ), экспрессирующие тенасцин С и ФРЭС-А, являются ключевыми медиаторами в процессе роста метастазов рака груди в легкие [134]. Прорастание метастазов меланомы в печень основывается на активации звездчатых клеток (резидентных печеночных фибробластов), участвующих в поддержании ангиогенеза [135]. При колоректальном раке ФАО, стимулированные TФР-β1, секретируют ИЛ-11 с целью повышения выживаемости колоректальных опухолевых клеток и успешности колонизации органа [136]. ФАО, стимулированные ТрФР, также усиливают интравазацию клеток колоректального рака и формирование отдаленных метастазов благодаря секреции станниокальцина-1 [137]. Фибронектин, ассоциированный с микроокружением легких, и клетки костного мозга, экспрессирующие рецепторы к ФРЭС-1, также поддерживают метастазирование клеток меланомы [138]. Утрата функции Fsp1 у мышей (мыши с нокаутом гена Fsp1) приводит к снижению подвижности фибробластов и ассоциировано с торможением темпов метастазирования [139]. ФАМ могут быть активированы в месте метастазирования в результате распространения злокачественных клеток или воспалительного ответа. Они могут появиться из других частей опухоли или из костного мозга и сразу вовлекаться в метастатический процесс; также они могут происходить из резидентных тканевых фибробластов. Их различное происхождение может также объяснять их функциональную гетерогенность.
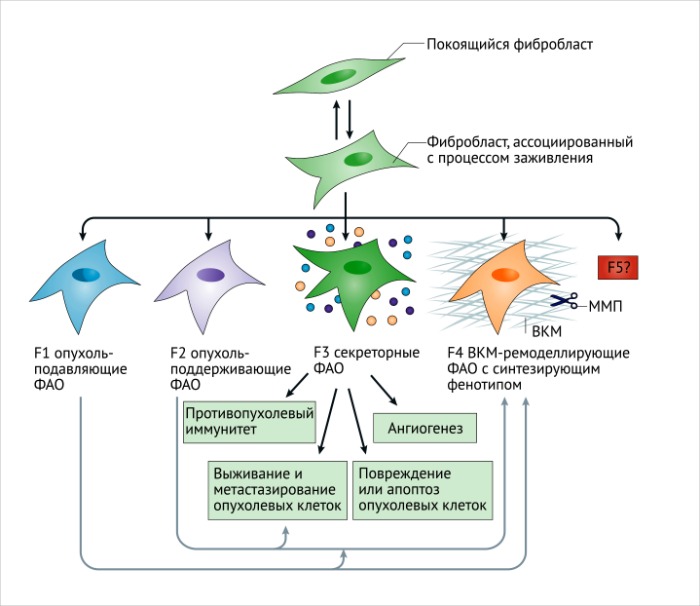
ФАО и опухолевый метаболизм
Клеточный метаболизм ФАО схож с метаболизмом быстро пролиферирующих клеток, который основывается на аэробном гликолизе [140]. Виновниками метаболических сдвигов при активации фибробластов являются TФР-β, ТрФР, гипоксия, индуцируемый гипоксией фактор 1α (HIF1α) и супрессия кавеолина-1 активными формами кислорода (АФК) [ССЫЛКИ 98, 141, 142].
Метаболическая адаптация может отображать ответ ФАО на внутриопухолевую гипоксию, вероятнее всего, обусловленную попыткой поддержания пролиферативной программы (РИС. 6).
Усиленный эффект Варбурга в ФАО связан с повышенной катаболической активностью и аутофагией [98, 141, 143].
Такие метаболические адаптации играют важнейшую роль в перенаправлении нутриентов на нужды других клеток ОМО и злокачественных клеток. В частности, повышенная продукция лактата в фибробластах с нокаутированным геном кавеолина-1, так же как и увеличенный уровень кетоновых тел, жирных кислот и глутамина наблюдаются в связи с их возможным участием в роли “топлива” для митохондриального дыхания в анаболических злокачественных клетках [141, 144]. ФАО могут принимать участие в анаболическом и катаболическом балансе опухолевых клеток в ОМО, представленном местами в достаточно разнородном виде. Повышенная активность анаболических процессов и митохондриального дыхания у злокачественных клеток может привести к усилению резистентности к терапевтическим средствам и началу метастазирования 145.
Метаболическое перепрограммирование ФАО зависит не только от паракринных влияний злокачественных клеток, но и от внутриклеточных контактов между ними, которые способны стимулировать взаимное метаболическое репрограммирование [148]. Исследования по установлению полного профиля экспрессируемых белков у ФАО выявило подавление изоцитратдегидрогеназы 3А (ИДГ-3А), стабилизацию HIF1α и индукцию генов гликолиза в условиях нормального содержания кислорода [142]. Активация р62 в ФАО ведет к ослаблению клеточного окислительно-восстановительного потенциала и повышению уровня ИЛ-6, обусловленного АФК [149]. Несмотря на то что специфические механизмы, инициирующие репрограммирование метаболизма в ФАО, требуют дальнейших исследований, уже становится очевидным, что метаболические изменения, однажды возникнув, получают способность к самоподдержанию – в частности, вероятно, благодаря эпигенетическим перестройкам. Однако метаболический симбиоз ФАО и злокачественных клеток может динамически развиваться лишь в ответ на активность кислорода, доступность внеклеточных метаболитов и сигнальные пути хемокинов и цитокинов. Гаплонедостаточность “датчика” кислорода и регулятора HIF1α пролилгидроксилазы 2 (PHD2; также известного как EGLN1) в MMTV-PyMT (средний Т-антиген полиомавируса мышиной опухоли молочной железы) у мышей снижает активацию ФАО in vivo и ассоциирована со снижением интенсивности метастазирования в легкие, вероятно, благодаря уменьшению продукции ТФР-β1 и репрограммированию ФАО [150]. Гипоксия также может играть прямую роль в активации ФАО [150]. Эти наблюдения говорят о том, что рецепция кислорода и метаболическое репрограммирование ФАО может влиять на их функцию (или функции) в растущих опухолях. Наконец, метаболизм ФАО может адаптироваться в ответ на курсы осуществляемой противоопухолевой терапии (о чем подробнее обсуждается ниже). Вклад изменений в метаболических программах ФАО, в основном, был изучен в контексте их влияния на рост злокачественных клеток, пролиферацию и инвазивность. В ходе метаболизма ФАО из-за нарушений в доступности метаболитов могут также индуцироваться проопухолевые биохимические циклы. Недостаточное поступление триптофана и аргинина в организм играет ключевую роль в активации Т-клеток и лимфоцитов, а метаболическая адаптация ФАО может регулировать биодоступность этих воспалительных метаболитов 152. Цитотоксические CD8-экспрессирующие Т-клетки имеют гиперактивированный гликолитический метаболизм, а роль ФАО в обеспечении биодоступности нутриентов для иммуных клеток также подтверждает их иммуномодуляторную функцию. В этом отношении, метаболическая конкуренция между стромальными клетками может привести к снижению реактивности Т-клеток в опухолях [152]. Эпителиальный канцерогенез может запускать метаболическое перепрограммирование ФАО, а также их гликолитическую и катаболическую адаптации в целях обеспечения реципрокного метаболического симбиоза между ФАО и злокачественными клетками [156]. Произвольные или специфически обусловленные симбиотические отношения между ФАО и опухолевыми клетками могут быть потенциальной мишенью для противоопухолевой терапии. Однако являются ли такие симбиотические отношения причиной развития метастазов – в целом, остается неизвестным.
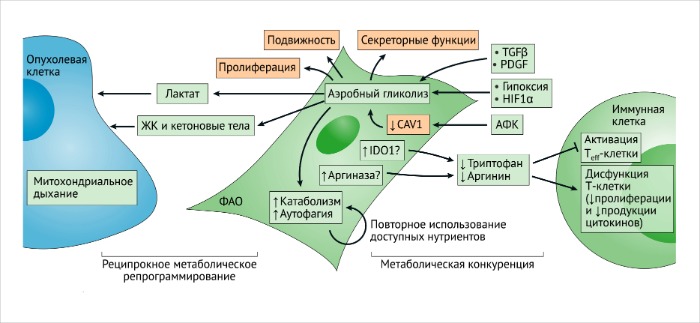
ФАО и противоопухолевый иммунный ответ
Плейотропное иммуномодуляторное влияние ФАО может быть прямым и непрямым. ФАО могут приобретать секреторный фенотип, осуществляя синтез белков ВКМ, экспрессию ВКМ-преобразующих ферментов и синтез множества цитокинов и хемокинов. Секретом ФАО не только постоянно поддерживает их в активированном состоянии в период опухолевой прогрессии, но также может динамически изменяться по мере развития опухоли, таким образом, по-разному влияя на противоопухолевый иммунитет на разных стадиях злокачественного роста. Секретом этих клеток также способен действовать на регуляцию противоопухолевого иммунного ответа напрямую (РИС. 3). Несмотря на то что недавние исследования секретома подтвердили гипотезу о положительном влиянии ФАО-специфического секретома на активацию опухолевых иммунных клеток, подобные кинетические исследования имеют ограничения в отношении клеток, развивающихся in vitro 157, и поэтому, в целях достоверного изучения роли секретома ФАО в обеспечении иммуномодуляторного ответа, проводятся in vivo. Тем не менее, данные работы указывают на гетерогенность секретома фибробластов [157, 159] и его избирательное влияние на биологию опухоли с применением химиотерапии или без нее [160]. Дальнейшие исследования с использованием специфических методов удаления из фибробластов цитокинов и хемокинов в доклинических моделях опухолей раскроют более точный список иммуномодуляторных цитокинов и хемокинов, синтезируемых ФАО. В целом, ФАО рассматривают как активаторов иммуносупрессивного влияния ОМО [161, 162]. Однако, возможно, это больше зависит от условий, нежели является специфической особенностью ФАО. В условиях гипоксии в опухолевом микроокружении ФАО, злокачественные клетки, эндотелиоциты и иммунные клетки динамично взаимодействуют, что значительно усложняет систему их паракринного влияния друг на друга.
Более того, большинство исследований, направленных на изучение иммуномодуляторного влияния секретома ФАО и иммунного ответа основывается на данных, полученных in vitro или на исследованиях in vivo, при которых ФАО, размноженные до инъекции in vitro, физически взаимодействовали со злокачественными клетками in vivo. Секреция ФАО в определенных опухолях цитокинов, хемокинов и про-ангиогенных факторов, включая, например, ИЛ-6, ИЛ-4, ИЛ-8, ИЛ-10, фактор некроза опухолей (ФНО), TФР-β, CCL2, CCL5 (также известный как RANTES), CXCL9, CXCL10, SDF1, простагландин E2 (ПГ-E2), оксид азота (NO), ГФР и человеческий лейкоцитарный антиген G (HLAG), может иметь прямой и/или непрямой эффекты на противоопухолевый иммунитет 165. Хотя сигнальный путь ИЛ-6 и так связан с ограничением созревания дендритных клеток (ДК), что обуславливает снижение активации Т-клеток и индуцирует Т-клеточную толерантность, секретируемые фибробластами ИЛ-6, помимо прочего, перенаправляют моноциты на дифференцировку в макрофаги, блокируя их путь дифференцировки в ДК [167], и активирует тучные клетки [168]. До сих пор остается неизвестным, является ли ИЛ-6, продуцируемый ФАО, скорость-лимитирующим фактором для иммунных реакций, или же другие источники ИЛ-6 в ОМО и другие факторы, такие как TФР-β, также задействованы. TФР-β регулирует бесчисленное множество иммуносупрессивных реакций [169], и недавние исследования показали, что TФР-β участвует в дифференцировке Т-хелперов линии 17 [170], что делает иммуномодуляторные функции продуцируемых ФАО молекул ТФР-β еще сложнее [171]. На основе анализа данных, полученных in vitro, продукция ИЛ-4, ИЛ-6 и ИЛ-8 может индуцировать дифференцировку иммуносупрессивных миелоидных клеток [19, 172]. Секретируемые ФАО факторы CXCL14 также влияют на противоопухолевый иммунитет, в частности, индуцируя мобилизацию макрофагов в опухоли [173]. Эти исследования подчеркивают роль паракринной сигнализации между ФАО и иммунными клетками, которая контролирует процесс неоплазии и злокачественного роста [174, 175]. CCL2 также играет роль в прогрессии рака груди на доклинических моделях [176], но изучение его эффектов ограничено исследованиями in vitro [176, 177]. CCL2 продуцируется также макрофагами и ДК, а секретом ФАО может активировать его синтез в других стромальных клетках. Он может усиливать влияние ФАО на ОМО. Также было обнаружено, что в активации Т-клеток участвуют такие цитокины, как CXCL9, CXCL10 и SDF1, входящие в секретом ФАО (ССЫЛКА 178).
Было установлено, что культивированные фибробласты из здорового кишечника человека экспрессируют негативные регуляторные иммунные сигналы (например, лиганды программируемой клеточной смерти 1 и 2 – соответственно, PDL1 и PDL2) c потенциальным тормозным воздействием на активацию Т-клеток [179]. В группе ФАО, полученных от пациентов с раком легкого, экспрессия PDL1 и PDL2 может иметь иммуносупрессивный эффект на активацию Т-клеток ex vivo [180]. Остается неизвестным, участвуют ли ФАО в иммуносупрессивном адаптивном ответе на солидную опухоль благодаря экспрессии ко-регуляторных сигналов in vivo. ФАО также могут усиливать противоопухолевый иммунитет косвенно, благодаря воздействию на опухолевый ангиогенез (регулируя трансэндотелиальную миграцию иммунных клеток), а также благодаря синтезу молекул адгезии (например, внутриклеточной молекулы адгезии 1 – ICAM1), что обеспечивает иммунные клетки платформой для специфических реакций, приводящих к их активации, репрессии или поляризации [21]. Иммунные клетки также влияют на ФАО посредством паракринных сигналов, хотя вся динамическая система отношений между ФАО и клетками иммунной системы остается, по большей мере, неизученной. Поляризация фенотипа ФАО [181] в процессе опухолевого развития может обеспечивать более динамичное и гетерогенное влияние на противоопухолевый иммунитет (РИС. 5). ФАО и злокачественные клетки создают сеть белков ВКМ, которые ограничивают доступ иммунных клеток к опухолевым благодаря образованию физического барьера.
С другой стороны, преобразование ВКМ, отчасти регулируемое ФАО, может обеспечить контакт Т-клеток с опухолевыми [182]. Ремоделирование ВКМ высвобождает провоспалительные факторы роста и цитокины и открывает скрытые связывающие сайты, которые могут обеспечивать адгезию иммунных клеток [5, 183]. В генетически модифицированных животных моделях и исследованиях ортотопических трансплантаций опухолей в иммунокомпетентных мышах, таргетируя БАФ-экспрессирующие ФАО, был продемонстрирован противоопухолевый эффект посредством активации CD8-позитивных T-клеток и опосредуемого ими киллинга [184, 185]. Воздействие на БАФ-экспрессирующие ФАО с помощью ДНК-вакцины или терапия мышей траниластом для подавления синтеза коллагена (несмотря на его неспецифическое связывание с ФАО) приводили к мобилизации и активации CD8-позитивных T-клеток и иммунному контролю опухолевого роста [186, 187]. Общая картина протуморогенной роли ФАО еще остается под вопросом и некоторые данные свидетельствуют о том, что ФАО могут ограничивать развитие злокачественной опухоли [96, 188]. Специфическое снижение числа α-ГMA-позитивных ФАО у трансгенных мышей, экспрессирующих вирусную тимидинкиназу под контролем промотера Acta2 (кодирующего α-ГMA) и обладающих ганцикловир-регулируемой пролиферацией α-ГMA-позитивных клеток, приводило к развитию инвазивных опухолей, связанных с иммуносупрессивным адаптивным ответом и увеличенной инфильтрацией регуляторных Т-клеток [96]. Несмотря на то, что общая картина роли ФАО в активации роста опухоли основывается на экспериментах с ко-культивированием клеток, настоящие и будущие исследования позволят определить функциональную роль ФАО в прогрессии опухолей и модуляции противоопухолевого иммунитета.
ФАО и лекарственная резистентность
Трудноразрешимой клинической задачей в терапии злокачественных опухолей является развитие резистентности, которая возобновляет развитие опухоли и метастазов, несмотря на лечение. Резистентность опухоли к терапии определяется прогрессией опухолевых очагов, сопутствующей первичному ответу на терапевтическое вмешательство или развившейся по отношению к нему вторично. Ранние исследования указывали на роль органоспецифического микроокружения в появлении резистентности [189], и ФАО считались ключевыми участниками прогрессии злокачественной опухоли при противоопухолевой терапии. Профиль генной экспрессии компонентов стромы ассоциирован с химиорезистентностью ( к 5-фторурацилу, эпирубицину и циклофосфамиду) при раке груди, что легко может позволить спрогнозировать ответ на химиотерапию при неоадьювантной терапии [190]. Несмотря на всю интригу, такие исследования, устанавливая связь между реактивной стромой и слабым ответом на химиотерапию, указывают, что ФАО не несут всей вины за появление резистентности к терапии, играя в этом лишь вспомогательную роль. Продолжающиеся попытки установить такие механические связи предпринимаются и являются предметом большого интереса для лучшего понимания противоопухолевой терапии. Механизмы развития резистентности, связанные со стромой, включают модуляцию путей, осуществляющих взаимодействие злокачественных клеток с клетками ВКМ, их адгезию и сигнальных путей цитокинов и хемокинов [191, 192]. ФАО также могут участвовать в повышенном внутриопухолевом давлении в интерстиции, таким образом косвенно ослабляя действие антибластомных препаратов [193]. Другие исследования предположили, что иммунная модуляция, проангиогенные процессы и метаболическое перепрограммирование ОМО, опосредованные ФАО, могут помочь в выживании злокачественных клеток и способствовать их уходу из-под контроля организма [112, 194, 195]. Хотя многие исследования на эту тему основываются на данных анализов, полученных in vitro и на моделях ксенотрансплантатов прогрессии опухоли, исследования ГМ-мышей и клинических образцов могут пролить свет на неизвестные функции ФАО в качестве ингибиторов противоопухолевой терапии. Повышенная адгезия злокачественных клеток к ВКМ может обеспечивать образование сигнальной платформы, которая усиливает механизмы выживания опухоли 196. Такие механизмы могут задействовать “спящий фенотип”, формирующийся благодаря остановке клеточного цикла при участии β1-интегрина [198, 199].
ФАО в меланомах с мутацией BRAF могут принимать участие в сопротивлении ингбированию BRAF путем образования богатого фибронектином ВКМ, усиливающего активацию ERK, опосредованную β1-интегрин-индуцированной киназой фокальных контактов (КФК) и SRC [200].
Наиболее важно то, что такие регулируемые ФАО программы не всегда повышают резистентность, но вместо этого могут создавать новые механизмы развития злокачественной опухоли, как, например, при разрастании резистентных клонов. Также важно заметить, что адгезия опухолевых клеток прямо к ФАО может также увеличивать лекарственную резистентность, вероятно, через N-кадгериновое гомотипичное связывание и усиление AKТ- сигналинга, направленного на выживание опухолевых клеток [201, 202]. Интересно, что приобретение адгезивных свойств опухолевыми клетками и их присоединение к ремоделированному ФАО внеклеточному матриксу может индуцировать ЭМП в опухолевых клетках, приводящий к терапевтической резистентности 205. Программа эпителиально-мезенхимального перехода может обеспечивать приобретение свойств химиорезистентности у опухолевых клеток благодаря индукции остановки клеточного цикла [96, 145] или изменению экспрессии клеточных транспортеров, обеспечивающих угнетение эффектов химиотерапии [206]. ФАО также могут обеспечивать резистентность к противоопухолевой лекарственной терапии с помощью растворимых факторов [191]. В этом отношении TФР-β, ИЛ-6 и ГФР, продуцируемые ФАО, являются потенциальными регуляторами лекарственной резистентности. TФР-β способен индуцировать мезенхимальные программы в опухолевых клетках, обеспечивая их адгезию к ВКМ, а ИЛ-6 индуцирует хорошо изученные сигнальные каскады, обеспечивающие выживание клеток [207]. ГФР также является ключевым модулятором ФАО-опосредованной резистентности, взаимодействуя с ингибиторами рецептора тирозинкиназы [208]. Было установлено, что ГФР, полученный из ФАО, усиливает резистентность у доклинических моделей опухолей при их лечении BRAF-V600E [209] или ингибиторами рецепторов к эпидермальному фактору роста (ЭФР) [210]. Злокачественные клетки, демонстрирующие высокую степень выживаемости после таргетной терапии, могут проявиться в ОМО благодаря ФАО-опосредованному аутокринному и паракринному сигнальным путям. Эти наблюдения доказывают рациональность применения лекарственных средств одновременно против ФАО и злокачественных клеток, в ходе чего, вероятно, активируются индуцируемые стромой синтетические пути, приводящие к гибели клеток [211].
Тем не менее, предполагаемое воздействие на Hedhehog-сигналинг для подавления проопухолевой активности ФАО аденокарциноме протоков поджелудочной железы (АПЖ) [212] привело к провалу второй фазы клинических испытаний [213]. Последующие исследования с долговременной оценкой доклинических моделей и специфическими стратегиями модификации генома подтвердили противоопухолевую активность ФАО и Hedhehog-сигналинга у ГМ-мышей с АПЖ [96, 188]. Тщательная проверка функции секретома ФАО в развитии лекарственной резистентности служит необходимым условием для лучшего прогноза значимости таргетирования ФАО в клинике. Некоторые комбинаторные стратегии на данный момент апробируются в клинике для преодоления ФАО-опосредованной лекарственной резистентности [191]. Тщательный анализ стромального ответа и адаптаций к терапии позволит понять конвергентную роль сигналинга между ФАО и злокачественными клетками во время терапии. Одна из стратегий включает ферментативную дезактивацию ВКМ, образованного ФАО. Деградация гиалуроновой кислоты [213, 215] или антиангиогенная терапия [216, 217] способны повлиять на перестройку и нормализацию опухолевых сосудов, что может обеспечить более эффективную доставку химиотерапевтических средств к злокачественным клеткам в солидных опухолях или усилить иммунорегулируемый противоопухолевый ответ. Несмотря на перспективность этих подходов, все они исходят из того, что ФАО стимулируют развитие опухоли. Эта концепция остается под вопросом и нуждается в дальнейших, более точных исследованиях.
ФАО – негативные регуляторы злокачественного роста
На сегодняшний день существует масса литературы, которая поддерживает идею о положительном влиянии ФАО на рост опухоли. Наиболее вероятно, что роль фибробластов в развитии опухоли носит непреднамеренный и побочный характер, обусловленный их участием в стромальном ответе организма на повреждение ткани.
В этом отношении, НАФ могут угнетать опухолевый рост благодаря обратимости эффектов TФР-β и ГФР, продуцируемых ФАО [218]. Потеря контроля за сигналингом, запускаемым TФР-β, в НАФ может индуцировать внутриэпителиальную неоплазию простаты, что указывает на супрессивную функцию НАФ в возникновении опухоли [8]. То, каким образом НАФ предотвращают опухолевый рост, остается практически неизвестным. Некоторые исследования также указывают, что элиминация стареющих фибробластов дермы вследствие дермабразии приводит к повышению экспрессии ИФР-1 и корригирует неадекватный ответ на облучение ультрафиолетом-В и опухолеобразование, обнаруженное в стареющей коже [220]. Такие процессы направлены на защиту стареющих кератиноцитов от канцерогенного влияния ультрафиолетового излучения и перерождения в плоскоклеточный рак [220]. Вероятно, что НАФ сдерживают опухолевый рост, в сравнении с активированными фибробластами. Недавние исследования показали, что ФАО способны сдерживать развитие аденокарциномы поджелудочной железы (АПЖ) благодаря редукции фиброза и гипоксии [188]. Другие исследования демонстрируют, что прямая инактивация α-ГMA-позитивных ФАО уменьшает выраженность фиброза и выживание мышей с АПЖ [96]. Активированные фибробласты могут влиять на защиту организма благодаря стимуляции врожденного и адаптивного противоопухолевого иммунитета. Многие исследования показали, что ФАО продуцируют такие иммуномодуляторные цитокины, как ИЛ-10, ТФР-β, ФНО, γ-интерферон и ИЛ-6, и помогают в активации и поляризационном распределении макрофагов, Т-лимфоцитов и естественных киллеров [162-165, 171, 175, 181]. Исследования с использованием мышиных моделей, которые позволяют извлекать активированные фибробласты, преследуют цель определить специфическую роль ФАО в модуляции противоопухолевого иммунитета. В связи с этим, клинические исследования, в ходе которых определяют экспрессию коллагена 1 типа и маркеры ФАО, включая α-ГMA и БАФ, при анализе исхода заболеваний демонстрируют, что пациенты с десмоплазией могут иметь хороший прогноз и полную выживаемость при АПЖ, раке молочной железы и раке легкого [221, 222]. При раке груди профиль экспрессии генов стромы может представлять ценную прогностическую информацию [223]. Структура десмопластичских стромальных клеток может также быть специфически ассоциирована с корреляциями, установленными при анализе клинической картины. Например, образуемые ФАО кавеолин-1 и β-рецептор ТрФР независимо ассоциированы с неблагоприятным прогнозом развития злокачественной опухоли молочной железы. Такие исследования необходимы в будущем для дальнейшего понимания потенциальной роли в ограничении роста опухолевой стромы и ФАО в прогрессии злокачественной опухоли.
Собранные данные свидетельствуют о возможности воздействия на ФАО с целью ингибирования роста злокачественных клеток. Однако еще предстоит, к примеру, найти такой агент, который специфически связывался бы с ФАО и воздействовал на него таргетно. Терапия против ФАО, направленная либо на их полное уничтожение, либо на их репрограммирование обратно в сторону нормального, покоящегося фенотипа, таким образом, вселяет некоторые надежды. Недавние исследования показали, что кальцитриол способен репрограммировать звездчатые клетки поджелудочной железы, дающих начало ФАО при АПЖ, по-видимому, возвращая их к нормальному фенотипу, подразумевающим, в числе прочего, отложение ретиноевой кислоты [225].
Такие доклинические попытки могут быть легко апробированы в клинике. Недавнее одобрение использования пирфенидона для лечения идиопатического фиброза легкого направлено на активацию ФАФ и усиление их секреторных функций [226]. Это дает возможность применять данный препарат при стандартной химиотерапии для вовлечения в терапию ФАО и злокачественных клеток. ФАО также могут быть таргетированы антителами, которые инактивируют БАФ (например, сибротузумаб) [227]; исследования по этому вопросу уже проводятся. Направленная терапия против активно пролиферирующих злокачественных клеток может также влиять на ФАО; таким образом, и химио-, и лучевая терапия способны индуцировать протуморогенные или противоопухолевые эффекты ФАО. Несмотря на то что воздействие на ФАО может послужить мощным противопухолевым терапевтическим средством, их функциональная гетерогенность и динамическая поляризация могут потребовать избирательного воздействия и более глубокого понимания ответных механизмов организма на терапию против ФАО.
Выводы и будущие перспективы
Становится очевидным, что покоящиеся фибробласты, вероятно, дают начало гетерогенной популяции активированных фибробластов. Различия в происхождении данных клеток и миофибробластов могут также быть основой для формирования гетерогенных популяций в опухолях. Мы считаем, что активированные фибробласты могут иметь подтипы, такие как F1 и F2 и так далее, похожие на подтипы макрофагов и других иммунных клеток (РИС, 5) [225]. Исследования по биологии развития, направленные на изучение различных подтипов мезенхимных клеток с различными функциями в различных тканях [51, 229, 230], могут выявить причины функциональной гетерогенности при патологиях, включая злокачественный рост. Уже на протяжении нескольких десятилетий активированные фибробласты рассматривались как соучастники злокачественных клеток в развитии опухоли. Однако эти клетки, вероятнее всего, непреднамеренно участвуют в канцерогенезе и могут служить как позитивными, так и негативными регуляторами опухолевого роста. Обладают ли различные подтипы фибробластов специфическими свойствами, или же популяции фибробластов функционируют вместе в зависимости от факторов среды на различных стадиях развития опухоли, остается неизвестным. Несмотря на это, изначальная функция фибробластов — это ответ на тканевое повреждение и индукция регенеративных процессов. В ответ на стимул, вырабатываемый поврежденными органами, и воспаление покоящиеся фибробласты активизируются, синтезируют факторы роста и регулируют увеличение ВКМ, а также влияют на воспаление и иммунитет.
Спорным является утверждение, что злокачественные клетки в среде, наполненной этими факторами, начинают заметно расти, мигрировать и вырабатывать механизмы выживания. Таким образом, фибробласты могут косвенно влиять на опухолевый рост. Первоначальный ответ нашего организма на злокачественный рост – это не противоборство, а контроль за тканевым повреждением.
Следующие 10 лет обещают подарить открытия многих скрытых тайн фибробластов.



