Нервная система рыб

Нервная система рыб гораздо примитивней нервной системы высших позвоночных и состоит из центральной (головной, спинной мозг), связанной с ней периферической (нервы, отходящие от головного и спинного мозга к органам) и вегетативной нервной системой (ганглии и нервы, иннервирующие мышцы внутренних органов и кровеносных сосудов сердца).
Функция мозга заключается в том, чтобы получать и интерпретировать электрические сигналы органов чувств — глаз, вкусовых почек, слуховых, осязательных органов и отвечать на них. Условно мозг состоит из трех сегментов: переднего мозга, среднего мозга и заднего мозга. Каждый сегмент связан с конкретными сенсорными входами.

В переднем мозгу происходит обработка информации, поступающей от органов обоняния. Эта часть регулирует движение и поведение рыбы. Например, передний мозг стимулирует и непосредственно участвует в регуляции икрометания, охраны икры, образования стаи, агрессии.
Промежуточный мозг отвечает за зрение рыбы: от него отходят зрительные нервы. К нижней стороне промежуточного мозга прилегает гипофиз, или питуитарная железа; в верхней части промежуточного мозга находится эпифиз, или пинеальная железа. Гипофиз и эпифиз — железы внутренней секреции. Промежуточный мозг участвует в координации движения, и работе других органов чувств.
Средний мозг имеет вид двух полушарий, а также самый большой объём. Доли (полушария) среднего мозга — первичные зрительные центры, обрабатывающие возбуждение, сигналы органов зрения, регуляции окраски, вкуса и равновесия; здесь же происходит и связь с мозжечком, продолговатым и спинным мозгом.
Мозжечок — маленький бугорок примыкающий сверху к продолговатому мозгу. Мозжечок отвечает за координацию движений, поддержание равновесия, мышечную деятельность. Он связан с рецепторами боковой линии, синхронизирует деятельность других отделов мозга.
Продолговатый мозг состоит из белого вещества и плавно переходит в спинной. Продолговатый мозг регулирует деятельность спинного мозга и вегетативной нервной системы. Важен для дыхательной, скелетно-мышечной, кровеносной и других систем рыбы. Если уничтожить эту часть мозга, например, перерезав рыбу в области за головой, то она быстро умирает. Кроме этого, продолговатый мозг отвечает за связь со спинным. От мозга отходят 10 пар черепно-мозговых нервов.
Хрящевые рыбы (акулы и скаты) имеют развитый передний мозг и обонятельные доли. Малоподвижные и донные рыбы — небольшой мозжечок и развитый передний и продолговатый отделы мозга, поскольку обоняние играет в их жизни значительную роль. У быстроплавающих рыб сильно развит средний мозг (зрительные доли) и мозжечок (координация движения). Слабые зрительные доли мозга у глубоководных рыб.

Спинной мозг — продолжение продолговатого мозга, его особенность — способность к быстрой регенерации и восстановлению деятельности при повреждении. Серое вещество в спинном мозге рыбы находится внутри, а белое — снаружи.
Спинной мозг — проводник и улавливатель рефлекторных сигналов. От спинного мозга отходят спинномозговые нервы, иннервирующие поверхность тела, туловищные мышцы, а через ганглии и внутренние органы. В спинном мозгу костистых рыб находится урогипофиз, клетки которого вырабатывают гормон, участвующий в водном обмене.
Вегетативная нервная система рыб — ганглии, находящиеся вдоль позвоночника. Клетки ганглиев связаны со спинномозговыми нервами и внутренними органами. Соединительные ветви ганглиев объединяют вегетативную нервную систему с центральной. Эти две системы независимы и взаимозаменяемы.
Проявление работы нервной системы рыбы — рефлекс. Если в конкретное время кормить рыб в одном и том же месте в пруду или в аквариуме, то гидрофаги будут скапливаться именно в этом месте к установленному часу. Кроме того, условные рефлексы у рыб вырабатываются на свет, форму, запах, звук, вкус, температуру воды.
Головной мозг рыб и его важнейшие отделы.




![]()
![]()
Головной мозг рыб очень мал, причем чем крупнее рыба, тем относительная масса мозга меньше. У крупных акул масса мозга составляет всего несколько тысячных долей процента массы тела. У осетровых и костистых рыб, весящих несколько килограммов, его масса достигает сотых долей процента массы тела. При массе рыбы в несколько десятков граммов мозг составляет доли процента, а у рыб, весящих меньше 1 г, мозг превышает 1 % массы тела. Это показывает, что рост мозга отстает от роста всего тела. Очевидно, основное развитие мозга происходит во время эмбрионально-личиночного развития. Разумеется, наблюдаются и межвидовые различия в относительной массе мозга.
Головной мозг состоит из пяти основных отделов: переднего, промежуточного, среднего, мозжечка и продолговатого мозга (СЛАЙД 6).
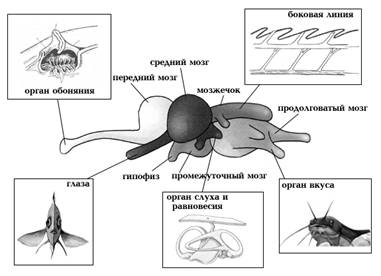
Строение головного мозга различных видов рыб различно и в большей степени зависит не от систематического положения рыб, а от их экологии. В зависимости от того, какие рецепторные аппараты преобладают у данной рыбы, соответственно развиваются и отделы мозга. При хорошо развитом обонянии увеличивается передний мозг, при хорошо развитом зрении — средний мозг, у хороших пловцов — мозжечок. У пелагических рыб хорошо развиты зрительные доли, относительно слабо развиты полосатые тела, хорошо развит мозжечок. У рыб, ведущих малоподвижный образ жизни, мозг характеризуется слабым развитием полосатых тел, маленьким шишкообразным мозжечком, иногда хорошо развитым продолговатым мозгом.
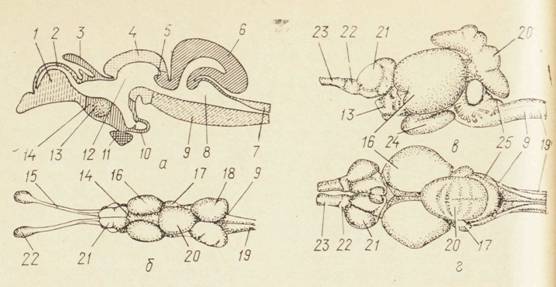
Рис. 14. Строение мозга костистых рыб:
а — схематическое изображение продольного разреза мозга; б — мозг карася, вид cut спины; в — мозг желтохвоста, вид сбоку; г — мозг желтохвоста, вид со спины; передний мозг; 2— первый мозговой желудочек; 3 — эпифиз; 4 — средний мозг; 5- заслонка мозжечка; 6 — мозжечок; 7 — мозговой канал; 8 — четвертый мозговой желудочек; 9 — продолговатый мозг; 10 — сосудистый мешок; 11 — гипофиз; 12 — третий мозговой желудочек; 13 — ядро зрительного нерва; 14 — промежуточный мозг; 15 — обонятельный тракт; 16 — зрительные доли; 11 — миндалевидные бугры; 18 — вагусные дилия 1У — спинной мозг; 20 — крыша мозжечка; 21 — обонятельные доли; 22 — обонятельная луковица; 23 — обонятельный тракт; 24 — гипоталамус; 25 — выступы мозжечка
Продолговатый мозг. Продолговатый мозг является продолжением спинного мозга. В своей передней части он переходит в задний отдел среднего мозга. Верхняя его часть — ромбовидная ямка — прикрыта эпендимой, на которой располагается заднее сосудистое сплетение. Продолговатый мозг выполняет ряд важных функций. Являясь продолжением спинного мозга, он играет роль проводника нервных импульсов между спинным мозгом и различными отделами головного мозга. Нервные импульсы проводятся как в нисходящем, т.е. к спинному мозгу, так и в восходящем направлениях — к среднему, промежуточному и переднему мозгу, а также к мозжечку.
В продолговатом мозге расположены ядра шести пар черепно-мозговых нервов (V—X). Из этих ядер, представляющих собой скопление нервных клеток, берут начало соответствующие черепно-мозговые нервы, выходяшие попарно с двух сторон мозга. Черепно-мозговые нервы иннервируют различные мышцы и рецепторные органы головы. Волокна блуждающего нерва иннервируют различные органы и боковую линию. Черепно-мозговые нервы могут быть трех видов: чувствительные, если в их составе имеются веточки, проводящие афферентные импульсы от органов чувств: двигательные, Hecyщие только эфферентную импульсацию к органам и мышцам; смешанные имеющие в своем составе чувствительные и двигательные волокна.
V пара — тройничный нерв. Начинается на боковой поверхности продол говатого мозга, делится на три ветви: глазничный нерв, который иннервирует переднюю часть головы; верхнечелюстной нерв, проходящий под глазом вдоль верхней челюсти и иннервирующий кожу передней части головы и нёбо; нижнечелюстной нерв, идущий вдоль нижней челюсти, иннервирующий кожу, слизистую оболочку ротовой полости и нижнечелюстную мускулатуру. Этот нерв содержит двигательные и чувствительные волокна..
VI пара отводящий нерв. Берет начало от дна продолговатого мозга, средней его линии, и иннервирует мышцы глаза,

и лабиринтовый аппарат. Его ядра располагаются между ядрами блуждающего нерва и основанием мозжечка.
IX – языкоглоточный нерв. Отходит от боковой стенки продолговатого
мозга и иннервирует слизистую оболочку нёба и мышцы первой жаберной дуги.
X – блуждающий нерв. Отходит от боковой стенки продолговатого мозга многочисленными веточками, которые образуют две ветви: боковой нерв, иннервирующий органы боковой линии в туловищной части; нерв жаберной крышки, иннервирующий жаберный аппарат и некоторые внутренние органы. По бокам ромбовидной ямки располагаются утолщения — вагальные доли, где расположены ядра блуждающего нерва.
У акул имеется XI нерв — конечный. Его ядра расположены на передней или нижней стороне обонятельных долей,, нервы проходят по дорзо-ла- теральной поверхности обонятельных трактов к обонятельным мешкам.
В области продолговатого мозга располагаются жизненно важные центры. Этот отдел мозга осуществляет регуляцию дыхания, сердечной деятельности, пищеварительного аппарата и др.
Дыхательный центр представлен группой нейронов, регулирующих дыхательные движения. Можно выделить центры вдоха и выдоха. Если разрушить половину продолговатого мозга, то дыхательные движения прекращаются только на соответствующей стороне. В области продолговатого мозга располагается также центр, регулирующий работу сердца и сосудов. Следующим важным центром продолговатого мозга является центр, регулирующий работу хроматофоров. При раздражении этого центра электрическим током происходит посветление всего тела рыбы. Здесь же находятся центры, регулирующие работу желудочно-кишечного тракта.
У рыб, имеющих электрические органы, разрастаются двигательные области продолговатого мозга, что приводит к образованию крупных электрических долей, которые являются своеобразным центром синхронизации разрядов отдельных электрических пластинок, иннервируемых различными мотонейронами спинного мозга.
У рыб, ведущих малоподвижный образ жизни, большое значение имеет вкусовой анализатор, в связи с чем у них развиваются специальные вкусовые доли.
В продолговатом мозге расположены в непосредственной близости от ядер VIII и X пар нервов — центры, ведающие движением плавников. При электрическом раздражении продолговатого мозга сзади от ядра X пары происходят изменения частоты и направления движения плавников.
Особое значение в составе продолговатого мозга имеет группа ганглиозных клеток в виде своеобразной нервной сети, называемой ретикулярной формацией. Она начинается в спинном мозге, далее встречается в продолговатом и среднем мозге.
У рыб ретикулярная формация связана с афферентными волокнами вестибулярного нерва (VIII) и нервов боковой линии (X), а также с волокнами, отходящими от среднего мозга и мозжечка. В ее составе находятся гигантские маунтнеровские клетки, иннервирующие плавательные движения рыб. Ретикулярная формация продолговатого, среднего и промежуточного мозга представляет единое в функциональном отношении образование, играющее важную роль в регуляции функций.
Регулирующее влияние на спинной мозг оказывает так называемая олива продолговатого мозга — ядро, хорошо выраженное у хрящевых рыб и хуже у костистых. Оно связано со спинным мозгом, мозжечком, промежуточным мозгом и участвует в регуляции движений.
У некоторых рыб, отличающихся высокой плавательной активностью, развивается добавочное ядро оливы, которое связано с деятельностью туловищной и хвостовой мускулатуры. Области ядер VIII и X пар нервов участвуют в перераспределении тонуса мышц и в осуществлении сложных координированных движений.
О важной роли среднего мозга рыб в отношении функций зрительного анализатора можно судить по выработке условных рефлексов на свет. Эти рефлексы у рыб можно выработать при удалении переднего, но при сохранении среднего мозга. При удалении среднего мозга условные рефлексы на свет исчезают, а выработанные ранее рефлексы на звук при этом не исчезают. После одностороннего удаления тектума у гольяна глаз рыбы, лежащий на противоположной стороне тела, слепнет, а при удалении тектума с двух сторон наступает полная слепота. Здесь же расположен центр зрительного хватательного рефлекса. Этот рефлекс заключается в том, что движения глаз, головы, всего туловища, вызываемые из области среднего мозга, надавлены на то, чтобы максимально способствовать фиксации объекта в области наибольшей остроты зрения — центральной ямки сетчатки. При электрической стимуляции определенных участков тектума форели появляются согласованные движения обоих глаз, плавников, муксулатуры тела.
Средний мозг играет важную роль в регуляции окраски рыб. При удалении глаз у рыб наблюдается резкое потемнение тела, а после двустороннего удаления тектума тело рыб светлеет.
В области тегментума располагаются ядра III и IV пар нервов, которые иннервируют мышцы глаз, а также вегетативные ядра, от которых отходят нервные волокна, иннервирующие мышцы, меняющие ширину зрачка.
Тектум тесно связан с мозжечком, гипоталамусом и через них—с передним мозгом. Тектум у рыб является одной из важнейших систем интеграции, он координирует функции соматосенсорной, обонятельной и зрительной систем. Тегментум связан с VIII парой нервов (акустические) и с рецепторным аппаратом лабиринтов, а также с V парой нервов (тройничные). К ядрам среднего мозга подходят афферентные волокна от органов боковой линии, от слухового и тройничного нервов. Все эти связи среднего мозга обеспечивают исключительную роль этого отдела центральной нервной системы у рыб в нервно-рефлекторной деятельности, имеющей приспособительное значение. Тектум у рыб, по-видимому, является основным органом замыкания временных связей.
Роль среднего мозга не ограничивается его связью со зрительным анализатором. В тектуме найдены окончания афферентных волокон от обонятельных и вкусовых рецепторов. Средний мозг рыб является ведущим центром регуляции движения. В области тегментума у рыб расположен гомолог красного ядра млекопитающих, функция которого заключается в регуляции тонуса мышц.
При повреждении зрительных долей падает тонус плавников. При удалении тектума с одной стороны возрастает тонус разгибателей противоположной стороны и сгибателей на стороне операции — рыба изгибается в сторону операции, начинаются манежные движения (движения по кругу). Это говорит о значении среднего мозга в перераспределении тонуса антагонистических мышц. При разобщении среднего и продолговатого мозга появляется повышенная спонтанная активность плавников. Из этого следует, что средний мозг оказывает тормозящее влияние на центры продолговатого и спинного мозга.
Промежуточный мозг. Промежуточный мозг состоит из трех образований: эпиталамуса — самой верхней надбугорной области; таламуса — средней части, содержащей зрительные бугры и гипоталамуса — подбугорной области. Этот отдел мозга у рыб частично прикрыт крышей среднего мозга.
Эпиталамус состоит из эпифиза или пинеального органа и габенулярных ядер.
Эпифиз — рудимент теменного глаза, он функционирует в основном как эндокринная железа. К эпиталамусу относится также уздечка (габенула), расположенная между передним мозгом и крышей среднего мозга. Она представлена двумя габенулярными ядрами, соединенными особойй связкой, к которым подходят волокна от эпифиза и обонятельные волокна переднего мозга. Таким образом, эти ядра имеют отношение к световосприятию и обонянию.
Эфферентные волокна идут к среднему мозгу и к нижерасположенным центрам. Зрительные бугры расположены в центральной части промежуточного мозга, своими внутренними боковыми стенками они ограничивают третий желудочек.
В таламусе различают дорзальную и вентральную области. В дорзальном таламусе у акул выделяют ряд ядер: наружное коленчатое тело, переднее, внутреннее и медиальное ядра.
Ядра зрительных бугров являются местом дифференцирования восприятий различных видов чувствительности. Сюда поступают афферентные влияния от различных органов чувств, здесь же происходят анализ и синтез афферентной сигнализации. Таким образом, зрительные бугры являются органом интеграции и регуляции чувствительности организма, а также принимают участие в осуществлении двигательных реакций. При разрушении промежуточного мозга у акул наблюдались исчезновение спонтанных движений, а также нарушение координации движений.
В состав гипоталамуса входит непарный полый выступ — воронка, которая образует особый, оплетенный сосудами орган — сосудистый мешок.
По бокам сосудистого мешка располагаются его нижние доли. У слепых рыб они очень малы. Считают, что эти доли связаны со зрением, хотя есть предположения, что этот отдел мозга связан с вкуосовыми окончаниями.
Сосудистый мешок хорошо развит у глубоководных морских рыб. Стенки его выстланы мерцательным кубическим эпителием, здесь же расположены нервные клетки, которые называются рецепторами глубины. Считают, что сосудистый мешок реагирует на изменения давления, а его рецепторы участвуют в регуляции плавучести; рецепторные клетки сосудистого мешка имеют отношение к восприятию скорости поступательного движения рыбы. Сосудистый мешок имеет нервные связи с мозжечком, благодаря этому сосудистый мешок участвует в регуляции равновесия и тонуса мускулатуры при активных движениях и колебаниях тела. У донных рыб сосудистый мешок рудиментарен.
Гипоталамус является главным центром, куда поступает информация из переднего мозга. Сюда поступают афферентные влияния от вкусовых окончаний и от акустико-латеральной системы. Эфферентные волокна от гипоталамуса идут к переднему мозгу, к дорзальному таламусу, тектуму, мозжечку, нейрогипофизу.
В гипоталамусе у рыб расположено преоптическое ядро, клетки которого обладают морфологическими признаками нервных клеток, но продуцируют нейросекрет.
Мозжечок. Он расположен в задней части головного мозга, частично прикрывает сверху продолговатый мозг. Различают среднюю часть — тело мозжечка — и два боковых отдела — ушки мозжечка. Передний конец мозжечка вдается в третий желудочек, образуя заслонку мозжечка.
У донных и малоподвижных рыб (удильщики, скорпены) мозжечокчок развит хуже, чем у рыб с большой подвижностью, Мозжечок у хищников (тунцы, скумбрии, тресковые), пелагических или планкктоноядных (харенгулы). У мормирид заслонка мозжечка гипертрофирована и иногда распространяется над мозальной поверхностью переднего мозга. У хрящевых рыб можно наблюдать увеличение поверхности мозжечка за счет образования складок.
У костистых рыб в задней, нижней части мозжечка находится скопление клеток под названием «латеральное ядро мозжечка», которое играет большую роль в поддержании тонуса мускулатуры.
При удалении у акулы половины аурикулярных долей тело ее начинает резко изгибаться в сторону операции (опистотонус). При удалении тела мозжечка с сохранением аурикулярных долей наступает нарушение тонуса мускулатуры и движения рыб только в том случае, если удаляется или перерезается нижняя часть мозжечка, где располагается латеральное ядро. При полном удалении мозжечка наступают падение тонуса (атония) и нарушение координации движений — рыбы плавают по кругу то в одну, то в другую сторону. Примерно через три недели утраченные функции восстанавливаются за счет регуляторных процессов других отделов мозга.
Удаление мозжечка у рыб, ведущих активный образ жизни (окуни, щуки и др.), вызывает сильные нарушения координации движений, сенсорные нарушения, полное исчезновение тактильной чувствительности, слабую реакцию на болевые раздражения.
Мозжечок у рыб, будучи связан через афферентные и эфферентные пути с тектумом, гипоталамусом, таламусом, продолговатым и спинным мозгом, может служить высшим органом интеграции нервной деятельности. После удаления тела мозжечка у поперечноротых и костистых рыб наблюдаются двигательные нарушения в виде качания тела из стороны в сторону. Если же удалить одновременно тело и заслонку мозжечка, то полностью нарушается двигательная деятельность, развиваются трофические нарушения и через 3— 4 недели животное гибнет. Это указывает на моторные и трофические функции мозжечка.
В ушки мозжечка поступают волокна от ядер VIII и X пар нервов. Ушки мозжечка достигают больших размеров у рыб, имеющих хорошо развитую баковую линию. Увеличение заслонки мозжечка также связано с развитием боковой линии. У золотых карасей выработанные дифференцировочные рефлексы на круг, треугольник и крест исчезали после коагуляции заслонки мозжечка и в последующем не восстанавливались. Это указывает на то, что мозжечок рыб является местом замыкания условных рефлексов, поступающих с органов боковой линии. С другой стороны, многочисленные эксперименты показывают, что у карпов с удаленным мозжечком в первые сутки после операции можно выработать двигательные и сердечные условные рефлексы на свет, звук и интероцептивные раздражения плавательного пузыря.
Передний мозг. Он состоит из двух частей. Дорзально лежит тонкая эпителиальная пластинка — мантия или плащ, отграничивающая от полости черепа общий желудочек; в основании переднего мозга лежат полосатые тела, которые с обе сторон соединяются передней связкой. Бока и крыша переднего мозга, образующие мантию, повторяют в общем форму лежащих под ними полосатых тел, от чего весь передний мозг кажется разделенным на два полушария, но истинного разделения на два полушария у костистых рыб не наблюдается.
В передней стенке переднего мозга развивается парное образование — обонятельные доли, которые иногда располагаются всей своей массой на передней стенке мозга, а иногда значительно вытягиваются в длину и нередко дифференцируются на основную часть (собственно обонятельная доля), стебелек и обонятельную луковицу.
У двоякодышащих рыб передняя стенка мозга вдвигается между полосатыми телами в виде складки, разделяющей передний мозг на два обособленных полушария.
В мантию поступают вторичные обонятельные волокна от обонятельной луковицы. Так как передний мозг у рыб представляет собой мозговую часть обонятельного аппарата, некоторые исследователи называют его обонятельным мозгом. После удаления переднего мозга наблюдается исчезновение выработанных условных рефлексов на обонятельные раздражители. После разобщения симметричных половин переднего мозга у карасей и карпов не наблюдается нарушений пространственного анализа зрительных и звуковых раздражителей, что указывает на примитивность функций этого отдела.
После удаления переднего мозга у рыб сохраняются условные рефлексы на свет, звук, магнитное поле, раздражения плавательного пузыря, раздраже ние боковой линии, вкусовые раздражители. Таким образом, дуги условных рефлексов на эти раздражители замыкаются на других уровнях головного мозга. Помимо обонятельных передний мозг рыб выполняет и некоторые другие функции. Удаление переднего мозга приводит к снижению двигательной активности у рыб.
Для разнообразных и сложных форм поведения рыб в стае необходима целостность переднего мозга. После его удаления рыбы плавают вне стаи. Вырабатывание условных рефлексов, наблюдающееся в условиях стаи, нарушается у рыб, лишенных переднего мозга. При удалении переднего мозга рыбы теряют инициативу. Так, нормальные рыбы, проплывая через частую решетку, избирают разные пути, а рыбы, лишенные переднего мозга, ограничиваются одним путем и обходят преграду с большим трудом. Интактные морские рыбы после 1—2-дневного пребывания в аквариуме не меняют своего поведения в море. Они возвращаются в стаю, занимают прежний охотничий участок, а если он занят, вступают в драку и изгоняют конкурента. Выпущенные в море оперированные особи не присоединяются к стае, не занимают свой охотничий участок и не закрепляют за собой нового, а если и удерживаются на ранее занимаемом, то не защищают его от конкурентов, хотя и не утрачивают способность защищаться. Если здоровые рыбы при возникновении опасной ситуации на своем участке умело используют особенности местности, последовательно перемещаются в одни и те же укрытия, то рыбам оперированные как бы забывают систему укрытий, используя случайные убежища.
Передний мозг играет важную роль и в половом поведении.
Удаление обеих долей у гемихромиса и сиамского петушка приводит к полной утрате полового поведения, у тиляпии нарушается способность к спариванию, у гуппи происходит задержка спаривания. У колюшки при удалении различных отделов переднего мозга изменяются (возрастают или уменьшаются) различ- функции — агрессивное, родительское или половое поведение. У самцов карася при разрушении переднего мозга пропадает половое влечение.
Таким образом, после удаления переднего мозга у рыб утрачиваются защитно-оборонительная реакция, способность заботиться о потомстве, способность к стайному плаванию, некоторые условные рефлексы, т.е. наблюдается изменение сложных форм условнорефлекторной деятельности и общеповеденческих безусловных реакций. Эти факты не дают исчерпывающего основания, что передний мозг у рыб приобретает значение органа интеграции, но позволяют предполагать, что он оказывает общее стимулирующее (тонизирующее) влияние на другие отделы мозга.



