За счет чего сокращаются мышцы
Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения.
1. Потенциал действия распространяется вдоль двигательного нервного волокна до его окончаний на мышечных волокнах.
2. Каждое нервное окончание секретирует небольшое количество нейромедиатора ацетилхолина.
3. Ацетилхолин действует на ограниченную область мембраны мышечного волокна, открывая многочисленные управляемые ацетилхолином каналы, проходящие сквозь белковые молекулы, встроенные в мембрану.
4. Открытие управляемых ацетилхолином каналов позволяет большому количеству ионов натрия диффундировать внутрь мышечного волокна, что ведет к возникновению на мембране потенциала действия.
5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна.
6. Потенциал действия деполяризует мышечную мембрану, и большая часть возникающего при этом электричества течет через центр мышечного волокна. Это ведет к выделению из саркоплазматического ретикулума большого количества ионов кальция, которые в нем хранятся.
7. Ионы кальция инициируют силы сцепления между актиновыми и миозиновыми нитями, вызывающие скольжение их относительно друг друга, что и составляет основу процесса сокращения мыщц.
8. Спустя долю секунды с помощью кальциевого насоса в мембране саркоплазматического ретикулума ионы кальция закачиваются обратно и сохраняются в ретикулуме до прихода нового потенциала действия. Удаление ионов кальция от миофибрилл ведет к прекращению мышечного сокращения.
Далее мы обсудим молекулярные механизмы этого процесса.
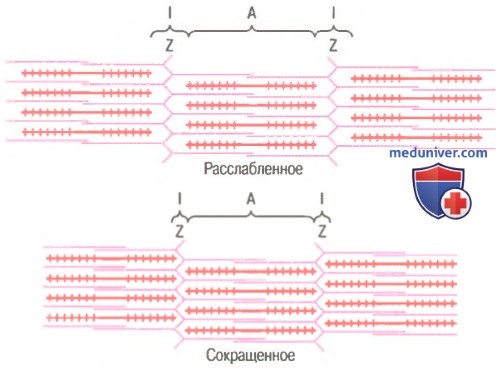 Миофибрилла в расслабленном и сокращенном состоянии. Показано (вверху), что актиновые нити (розовые) вдвинуты в пространства между миозиновыми нитями (красные). Сближение Z-дисков друг с другом (внизу).
Миофибрилла в расслабленном и сокращенном состоянии. Показано (вверху), что актиновые нити (розовые) вдвинуты в пространства между миозиновыми нитями (красные). Сближение Z-дисков друг с другом (внизу).
Молекулярные механизмы мышечного сокращения
Механизм скольжения нитей для мышечного сокращения. На рисунке показан основной механизм мышечного сокращения. Показано расслабленное состояние саркомера (вверху) и сокращенное состояние (внизу). В расслабленном состоянии концы актиновых нитей, отходящие от двух последовательных Z-дисков, лишь незначительно перекрываются. Наоборот, в сокращенном состоянии актиновые нити втягиваются внутрь между миозиновыми так сильно, что их концы максимально перекрывают друг друга. При этом Z-диски притягиваются актиновыми нитями к концам миозиновых. Таким образом, мышечное сокращение осуществляется путем механизма скольжения нитей.
Что заставляет нити актина скользить внутрь среди нитей миозина? Это связано с действием сил, генерируемых при взаимодействии поперечных мостиков, исходящих от нитей миозина, с нитями актина. В условиях покоя эти силы не проявляются, однако распространение потенциала действия вдоль мышечного волокна приводит к выделению из саркоплазматическо-го ретикулума большого количества ионов кальция, которые быстро окружают миофи-бриллы. В свою очередь, ионы кальция активируют силы взаимодействия между нитями актина и миозина, в результате начинается сокращение. Для осуществления процесса сокращения необходима энергия. Ее источником являются высокоэнергетические связи молекулы АТФ, которая разрушается до АДФ с высвобождением энергии. В следующих разделах мы приведем известные детали молекулярных процессов сокращения.
Молекулярные особенности сократительных нитей
Миозиновая нить. Она состоит из множества молекул миозина, молекулярная масса каждой составляет около 480000. На рисунке показана отдельная молекула; и также — объединение многих молекул миозина в миозиновую нить, а также взаимодействие одной стороны этой нити с концами двух актиновых нитей.
В состав молекулы миозина входят 6 полипептидных цепей: 2 тяжелые цепи с молекулярной массой около 200000 каждая и 4 легкие цепи с молекулярной массой около 20000 каждая. Две тяжелые цепи спирально закручиваются вокруг друг друга, формируя двойную спираль, которую называют миозиновым хвостом. С одного конца обе цепи изгибаются в противоположных направлениях, формируя глобулярную полипептидную структуру, называемую миозиновой головкой. Таким образом, на одном конце двойной спирали молекулы миозина образуются 2 свободные головки; 4 легкие цепи также включены в состав миозиновой головки (по 2 в каждой). Они помогают регулировать функцию головки во время мышечного сокращения.
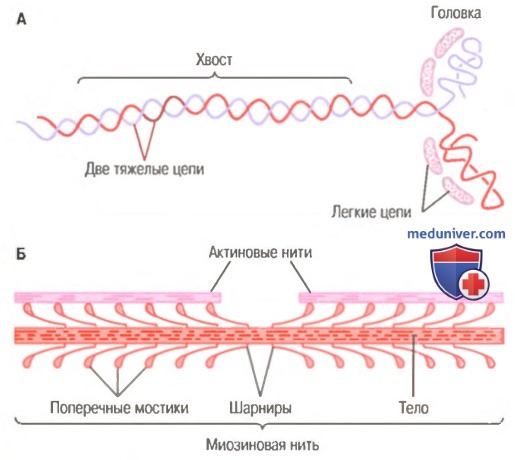 А. Молекула миозина.
А. Молекула миозина.
Б. Объединение многих молекул миозина в одну миозиновую нить.
Показаны также тысячи миозиновых поперечных мостиков и взаимодействие их головок с прилежащими актиновыми нитями.
Миозиновая нить состоит из 200 или более отдельных молекул миозина. Видно, что хвосты молекул миозина объединяются, формируя тело нити, а многочисленные головки молекул выдаются наружу по сторонам тела. Кроме того, наряду с головкой в сторону выступает часть хвоста каждой миозиновой молекулы, образуя плечОу которое выдвигает головку наружу от тела, как показано на рисунке. Выступающие плечи и головки вместе называют поперечными мостиками. Каждый поперечный мостик может сгибаться в двух точках, называемых шарнирами. Один из них расположен в месте, где плечо отходит от тела миозиновой нити, а другой — где головка крепится к плечу. Движение плеча позволяет головке или выдвигаться далеко наружу от тела миозиновой нити, или приближаться к телу. В свою очередь, повороты головки участвуют в процессе сокращения, что обсуждается в следующих разделах.
Общая длина каждой миозиновой нити остается постоянной и равна почти 1,6 мкм. В самом центре миозиновой нити на протяжении 0,2 мкм поперечных мостиков нет, поскольку снабженные шарнирами плечи отходят в стороны от центра.
Сама миозиновая нить сплетена таким образом, что каждая последующая пара поперечных мостиков смещена в продольном направлении относительно предыдущей на 120°, что обеспечивает распределение поперечных мостиков во всех направлениях вокруг нити.
АТФ-азная активность миозиновой головки. Есть и другая особенность миозиновой головки, необходимая для мышечного сокращения: миозиновая головка функционирует как фермент АТФ-аза. Как объясняется далее, это свойство позволяет головке расщеплять АТФ и использовать энергию расщепления высокоэнергетической связи для процесса сокращения.
Актиновая нить. Актиновая нить состоит из трех белковых компонентов: актина, тропомиозина и тропонина.
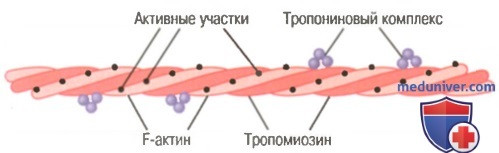 Актиновая нить, состоящая из двух спиралевидных цепочек молекул F-актина и двух цепочек молекул тропомиозина, расположенных в желобках между цепочками актина.
Актиновая нить, состоящая из двух спиралевидных цепочек молекул F-актина и двух цепочек молекул тропомиозина, расположенных в желобках между цепочками актина.
К одному концу каждой молекулы тропомиозина прикреплен тропониновый комплекс, который запускает сокращение.
Основой актиновой нити являются две цепи белковой молекулы F-актина. Обе цепи закручиваются в спираль так же, как и молекула миозина.
Каждая цепь двойной спирали F-актина состоит из полимеризованных молекул G-актина с молекулярной массой около 42000. К каждой молекуле G-актина прикреплена 1 молекула АДФ. Полагают, что эти молекулы АДФ являются активными участками на актиновых нитях, с которыми взаимодействуют поперечные мостики миозиновых нитей, обеспечивая мышечное сокращение. Активные участки на обеих цепях F-актина двойной спирали расположены со смещением таким образом, что вдоль всей поверхности актиновой нити встречается один активный участок примерно через каждые 2,7 нм.
Длина каждой актиновой нити — около 1 мкм. Основания актиновых нитей прочно встроены в Z-диски; концы этих нитей выступают в обоих направлениях, располагаясь в пространствах между миозиновыми молекулами.
Молекулы тропомиозина. Актиновая нить также содержит другой белок — тропомиозин. Каждая молекула тропомиозина имеет молекулярную массу 70000 и длину 40 нм. Эти молекулы спирально оплетают спираль из F-актина. В состоянии покоя молекулы тропомиозина располагаются поверх активных участков актиновых нитей, препятствуя их взаимодействию с миозиновыми нитями, лежащему в основе сокращения.
Тропонин и его роль в мышечном сокращении. По ходу молекул тропомиозина к ним периодически прикреплены другие белковые молекулы, называемые тропонином. Они представляют собой комплексы трех слабосвязанных белковых субъединиц, каждая из которых играет специфическую роль в регуляции мышечного сокращения. Одна из субъединиц (тропонин I) имеет высокое сродство к актину, другая (тропонин Т) — к тропомиозину, третья (тропонин С) — к ионам кальция. Считают, что этот комплекс прикрепляет тропомиозин к актину. Высокое сродство тропонина к ионам кальция, как полагают, инициирует процесс сокращения, о чем говорится в следующей статье.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Механизм мышечного сокращения
Содержание
Нервно-мышечная реакция на силовую тренировку [ править | править код ]
Источник: «Программы тренировок», научное изд.
Автор: профессор, доктор наук Тудор Бомпа, 2016 г.
Структура мышц [ править | править код ]
Помимо этого, способность мышцы сокращаться и прилагать силу зависит конкретно от ее вида, площади поперечного сечения, а также длины и количества волокон внутри мышцы. Число волокон определяется генетикой, и на него невозможно повлиять с помощью тренировок; однако тренировки в состоянии изменить другие переменные. Например, число и толщина миозиновых нитей увеличивается посредством упорных тренировок с максимальной силовой нагрузкой. Увеличение толщины мышечных нитей увеличивает размер мышцы и силу сокращений.
Разные двигательные единицы реагируют на разные нагрузки при тренировках. Например, выполнение жима лежа с 60% повторного максимума задействует определенную совокупность двигательных единиц, тогда как более крупные двигательные единицы ожидают более высокой нагрузки. Поскольку последовательное задействование двигательных единиц зависит от нагрузки, необходимо разрабатывать специальные программы, чтобы активизировать и адаптировать основные группы двигательных единиц и мышечных волокон, играющих доминирующую роль в избранном виде спорта. К примеру, в тренировках для спринта на короткую дистанцию и легкоатлетических дисциплин (таких как толкание ядра) следует использовать тяжелые нагрузки, чтобы способствовать развитию силы, необходимой для оптимизации скорости и взрывных действий.
Мышечные волокна выполняют разные биохимические (метаболические) функции; выражаясь конкретнее, одни лучше приспособлены с физиологической точки зрения к работе в анаэробных условиях, а другие лучше работают в аэробных условиях. Волокна, которые используют кислород для выработки энергии, называются аэробными, тип I, красными или медленными. Волокна, которым кислород не требуется, называются анаэробными, тип II, белыми или быстрыми. Быстрые мышечные волокна, в свою очередь, делятся на подтипы IIА и IIХ (иногда называемые IIВ, хотя у людей тип IIВ практически не встречается [1] ).
Медленные и быстрые волокна существуют примерно в равной пропорции. Однако в зависимости от их функций, в некоторых группах мышц (например, подколенные сухожилия, бицепсы) содержится больше быстрых волокон, тогда как в других (например, в камбаловидной мышце) содержится больше медленных волокон. В таблице 2.1 мы сравниваем характеристики быстрых и медленных волокон.
Сравнение быстрых и медленных волокон
Красные, тип I, аэробные
Белые, тип II, анаэробные
• Развивают долгие, продолжительные сокращения
• Применяются для развития выносливости
• Активизируются во время низко- и высокоинтенсивной деятельности
от 300 до 500 (или более) мышечных волокон
• Развивают короткие, сильные сокращения
• Применяются для развития скорости и силы
• Активизируются только во время высокоинтенсивной деятельности
Сокращение быстрой двигательной единицы более быстрое и мощное, чем сокращение медленной двигательной единицы. В результате пропорция быстрых волокон, как правило, выше в организме успешных спортсменов, занимающихся скоростно-силовыми видами спорта, но они также быстрее утомляются. Спортсмены с более высоким скоплением медленных волокон, напротив, обычно преуспевают в видах спорта на выносливость, поскольку они могут выполнять нагрузки низкой интенсивности в течение более продолжительного времени.
В распределении типов мышечных волокон у спортсменов, занимающихся разными видами спорта, могут наблюдаться различия. Это иллюстрируют рис. 2 и 2.3, представляющие общий процент содержания быстрых и медленных мышечных волокон у спортсменов в избранных видах спорта. Например, существенная разница между спринтерами и марафонцами четко дает понять, что успех в некоторых видах спорта хотя бы частично определяется генетическим составом мышечных волокон спортсмена.


Механизм мышечных сокращений [ править | править код ]
Первичные мышцы в первую очередь отвечают за суставное действие, которое является частью объемного силового движения или технической способности. Например, во время флексии локтя (сгибание бицепса) первичной мышцей является двуглавая мышца, тогда как трехглавая мышца (трицепс) выступает в роли антагониста и должна быть расслаблена, чтобы обеспечить беспрепятственное действие. В дополнение к этому стабилизаторы, или фиксаторы (обычно это меньшие мышцы), сокращаются изометрически, чтобы закрепить кость так, чтобы у первичных мышц была прочная база, откуда начинать натяжение. Мышцы других конечностей также могут принимать в этом участие, выступая в роли стабилизаторов, позволяющих первичным мышцам выполнять необходимые движения. Например, когда дзюдоист тянет соперника на себя, удерживая его за дзюдоги, мышцы его спины, ног и живота сокращаются изометрически, чтобы обеспечить стабильное основание для действия локтевых сгибателей (бицепсов), плечевых разгибателей (задние дельты) и лопаточных аддукторов и депрессоров (трапециевидная мышца и широчайшая мышца спины).
Механика мышечных сокращений [ править | править код ]
Если мышцу стимулировать коротким электрическим импульсом, спустя небольшой латентный период происходит ее сокращение. Такое сокращение называется «одиночное сокращение мышцы». Одиночное мышечное сокращение длится около 10-50 мс, причем оно достигает максимальной силы через 5-30 мс.
Каждое отдельное мышечное волокно подчиняется закону «все или ничего», т. е. при силе раздражения выше порогового уровня происходит полное сокращение с максимальной для данного волокна силой, а ступенчатое повышение силы сокращения по мере увеличения силы раздражения невозможно. Поскольку смешанная мышца состоит из множества волокон с различным уровнем чувствительности к возбуждению, сокращение всей мышцы может быть ступенчатым в зависимости от силы раздражения, при этом при сильных раздражениях происходит активация глубжележащих мышечных волокон.
Механизм скольжения филаментов [ править | править код ]

Укорочение мышцы происходит за счет укорочения образующих ее саркомеров, которые, в свою очередь, укорачиваются за счет скольжения относительно друг друга актиновых и миозиновых филаментов (а не укорочения самих белков). Теория скольжения филаментов была предложена учеными Huxley и Hanson (Huxley, 1974; рис. 1). (В 1954 г. две группы исследователей — X. Хаксли с Дж. Хэнсон и А. Хаксли с Р. Нидергерке — сформулировали теорию, объясняющую мышечное сокращение скольжением нитей. Независимо друг от друга они обнаружили, что длина диска А оставалась постоянной в расслабленном и укороченном саркомере. Это позволило предположить, что есть два набора нитей — актиновые и миозиновые, причем одни входят в промежутки между другими, и при изменении длины саркомера эти нити каким-то образом скользят друг по другу. Сейчас эта гипотеза принята почти всеми.)
Актин и миозин — два сократительных белка, которые способны вступать в химическое взаимодействие, приводящее к изменению их взаимного расположения в мышечной клетке. При этом цепочка миозина прикрепляется к актиновой нити с помощью целого ряда особых «головок», каждая из которых сидит на длинной пружинистой «шее». Когда происходит сцепление между миозиновой головкой и актиновой нитью, конформация комплекса этих двух белков изменяется, миозиновые цепочки продвигаются между актиновыми нитями и мышца в целом укорачивается (сокращается). Однако, чтобы химическая связь между головкой миозина и активной нитью образовалась, необходимо подготовить этот процесс, поскольку в спокойном (расслабленном) состоянии мышцы активные зоны белка актина заняты другим белком — тропохмиозином, который не позволяет актину вступить во взаимодействие с миозином. Именно для того, чтобы убрать тропомиозиновый «чехол» с актиновой нити, требуется быстрое выливание ионов кальция из цистерн саркоплазматического ретикулума, что происходит в результате прохождения через мембрану мышечной клетки потенциала действия. Кальций изменяет конформацию молекулы тропомиозина, в результате чего активные зоны молекулы актина открываются для присоединения головок миозина. Само это присоединение осуществляется с помощью так называемых водородных мостиков, которые очень прочно связывают две белковые молекулы — актин и миозин — и способны в таком связанном виде находиться очень долго.
Для отсоединения миозиновой головки от актина необходимо затратить энергию аденозинтрифосфа-та (АТФ), при этом миозин выступает в роли АТФазы (фермента, расщепляющего АТФ). Расщепление АТФ на аденозиндифосфат (АДФ) и неорганический фосфат (Ф) высвобождает энергию, разрушает связь между актином и миозином и возвращает головку миозина в исходное положение. В дальнейшем между актином и миозином могут снова образовываться поперечные связи.
При отсутствии АТФ актин-миозиновые связи не разрушаются. Это и является причиной трупного окоченения (rigor mortis) после смерти, т. к. останавливается выработка АТФ в организме — АТФ предотвращает мышечную ригидность.
Даже при мышечных сокращениях без видимого укорочения (изометрические сокращения, см. выше) активируется цикл формирования поперечных связей, мышца потребляет АТФ и выделяет тепло. Головка миозина многократно присоединяется на одно и то же место связывания актина, и вся система миофиламентов остается неподвижной.
Внимание: Сократительные элементы мышц актин и миозин сами по себе не способны к укорочению. Мышечное укорочение является следствием взаимного скольжения миофиламентов относительно друг друга (механизм скольжения филаментов).
Как же образование поперечных связей (водородных мостиков) переходит в движение? Одиночный саркомер за один цикл укорачивается приблизительно на 5-10 нм, т.е. примерно на 1 % своей общей длины. За счет быстрого повторения цикла поперечных связей возможно укорочение на 0,4 мкм, или 20% своей длины. Поскольку каждая миофибрилла состоит из множества саркомеров и во всех них одновременно (но не синхронно) образуются поперечные связи, суммарно их работа приводит к видимому укорочению всей мышцы. Передача силы этого укорочения происходит через Z-линии миофибрилл, а также концы сухожилий, прикрепленных к костям, в результате чего и возникает движение в суставах, через которые мышцы реализуют перемещение в пространстве частей тела или продвижение всего тела.
Связь между длиной саркомера и силой мышечных сокращений [ править | править код ]
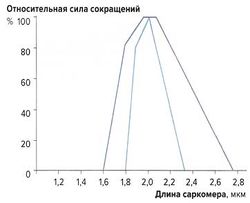
Наибольшую силу сокращений мышечные волокна развивают при длине 2-2,2 мкм. При сильном растяжении или укорочении саркомеров сила сокращений снижается (рис. 2). Эту зависимость можно объяснить механизмом скольжения филаментов: при указанной длине саркомеров наложение миозиновых и актиновых волокон оптимально; при большем укорочении миофиламенты перекрываются слишком сильно, а при растяжении наложение миофиламентов недостаточно для развития достаточной силы сокращений.
Скорость укорочения мышечных волокон [ править | править код ]
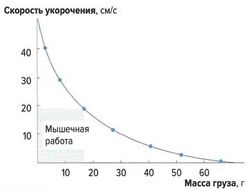
Скорость укорочения мышцы зависит от нагрузки на эту мышцу (закон Хилла, рис. 3). Она максимальна без нагрузки, а при максимальной нагрузке практически равна нулю, что соответствует изометрическому сокращению, при котором мышца развивает силу, не изменяя своей длины.
Влияние растяжения на силу сокращений: кривая растяжения в покое [ править | править код ]
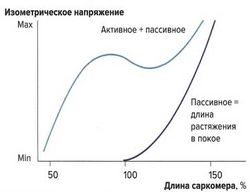
Важным фактором, влияющим на силу сокращений, является величина растяжения мышцы. Тяга за конец мышцы и натяжение мышечных волокон называются пассивным растяжением. Мышца обладает эластическими свойствами, однако в отличие от стальной пружины зависимость напряжения от растяжения не линейна, а образует дугообразную кривую. С увеличением растяжения повышается и напряжение мышцы, но до определенного максимума. Кривая, описывающая эти взаимоотношения, называется кривой растяжения в покое.
Данный физиологический механизм объясняется эластическими элементами мышцы — эластичностью сарколеммы и соединительной ткани, располагающимися параллельно сократительным мышечным волокнам.
Также при растяжении изменяется и наложение друг на друга миофиламентов, однако это не оказывает влияния на кривую растяжения, т. к. в покое не образуются поперечные связи между актином и миозином. Предварительное растяжение (пассивное растяжение) суммируется с силой изометрических сокращений (активная сила сокращений).
За счет чего сокращаются мышцы

На протяжении многих лет изучение процессов синтеза белков в скелетных мышцах при выполнении различных физических нагрузок остаётся актуальной проблемой биохимии и физиологии. Мышцы и их силовые характеристики очень важная составляющая организма каждого спортсмена, которая позволяет достигать результатов. В связи с прогрессивным развитием спорта и вовлечением большого количества людей в физическую культуру, тема здоровья спортсменов становится все более актуальной, интересной и увлекательной. Учитывая существующую сильную корреляцию между площадью поперечного сечения мышц и мышечной силой, стремление увеличить мышечную массу тела есть у каждого человека, занимающегося спортом. Кроме этого, необходимо помнить, что преобладание мышечной массы в организме благоприятно влияет на метаболические процессы.
Скелетная мышца – одна из наиболее пластичных структур в организме млекопитающих. При повышенной активности и нагрузке часто происходит увеличение её размеров, объёмов миофибриллярного аппарата, повышение сократительных возможностей (силы, мощности). Процесс прироста мышечной массы зависит от различных факторов: наследственных, конституциональных, а также пола, возраста, метаболизма, гормонального фона. Кроме того, с приобретением опыта тренировок становится все труднее увеличить мышечную массу, поэтому важно понимать и активно использовать все возможные механизмы этого процесса.
Клетки поперечно-полосатой мускулатуры отличаются от гладкомышечных миоцитов. Клетки скелетных мышц образуют многоядерный синцитий, основное вещество которого формируют миофибриллы, состоящие из толстых и тонких миофиламентов. Первый тип образуют молекулярные единицы и миозин, а второй тип содержит тропомиозин с тропонином и F-актин. Многие авторы считают скелетную мускулатуру гетерогенной системой относительно устройства и выполняемых функций, несмотря на её строгую организацию. Данное свойство помогает мышцам соответствовать возлагаемой на них функции. Так путём изменения количества саркомеров и миофибрилл обеспечивается их функциональная реорганизация [1].
Работа мышц проявляется их сокращением, которое начинается с появления очага возбуждения на нейромышечных окончаниях. Наружная мембрана деполяризуется, открываются кальциевые каналы, и концентрация кальция внутри клетки возрастает. Ионы кальция связываются с тропонином, при этом конформируется тропониновый комплекс. Участки цепей миозина связываются с актином, что сопровождается высвобождением энергии вследствие расщепления АТФ до АДФ и остатка фосфорной кислоты. Угол между лёгкой и тяжёлой цепями миозина изменяется и актиновый филамент перемещается к центру саркомера, что приводит к изменению длины мышцы, её сокращению [1, 2].
Клетки скелетных мышц подразделяются на два типа:
А) Миосателлиты – взрослые стволовые клетки мышечной ткани. Представляют собой основу для обновления мышц и прироста их массы;
Б) Миосимпласты – формируют многоядерный синцитий. Сами по себе являются мышечными тубами с миофибриллами внутри, по периферии которых располагаются ядра.
Нагрузки, оказываемые на мышцы, и само мышечное сокращение имеют некую зависимость. Предполагается, что первое будет напрямую соответствовать второму. Это достигается за счёт усиления экспрессии генов сократительных белков и энзимов обменных процессов. Мышечная активность сопровождается количественными и качественными изменениями в миоцитах того типа, которые необходимы для наиболее эффективного осуществления выполняемой работы [2].
Мышечные волокна делятся на медленные (I тип) и быстрые (II тип). Оба этих типа имеют различный состав, включающий в себя сократительные белки, ферменты энергетического обмена и внутриклеточный кальций.
Увеличение силы мышц проявляется структурными перестройками, которые затрагивают нервную и мышечные системы. Изменения в нервной системе проявляются трансформацией величины кортикальных полей, которые регулируют выполнение определённого вида движения, влиянием на синхронизацию моторных единиц и на обучение определенных мышц, отвечающих за выполнение данного вида движений. Таким образом, наибольшая активность мышц наблюдается именно тогда, когда она необходима для достижения максимального эффекта (активность мышц агонистов при одновременной пассивности антагонистов). Также наблюдается изменение частоты и устойчивости генерируемых импульсов и порога возбудимости мотонейронов. Изменения в мышечной системе могут быть связаны с гипертрофией скелетных мышц (увеличение размеров мышечного волокна) и с их гиперплазией (увеличение количества миоцитов) [3].
Но прежде чем переходить к последним двум процессам, необходимо разобраться с изменениями, происходящими в самих мышцах. В момент выполнения работы миоцит подвергается действию физических и гуморальных факторов (пассивные механические силы, гипоксемия, факторы роста, и т.д.). Они являются причиной запуска путей передачи сигнала внутри клеток, опосредуя транскрипцию и трансляцию генов, ответственных за синтез белков [2]. Изменения данных путей сопровождаются реорганизацией мышечных волокон, точнее их типов.
Одним из основных исходных сигналов является повышенная концентрация кальция внутри клетки и кальцинейрина. Кальцинейрин дефосфорилирует факторы транскрипции – NFAT (nuclear factor of activated T-cells), которые находятся в фосфорилированном состоянии [4]. Данные факторы в дефосфорилированной форме активируют гены-мишени, что способствует перестроению быстрых волокон в медленные.
По мере приспособления мышц к нагрузкам изменяются и процессы метаболизма в них. Существуют различные параметры, влияющие на формирование адаптивных механизмов в миоцитах при выполнении работы. Важнейшим является гипоксия, которая, в свою очередь активирует ферментные системы (фумараза, цитратсинтаза, ЛДГ) и запускает работу факторов транскрипции (PGC1). При недостатке кислорода происходит активация одной изоформы семейства гипоксия-индуцированных факторов (HIF; hypoxia inducible factor), которая проникает в ядро, связывается с определенным участком ДНК и активирует гены, отвечающие за гликолиз, потребление кислорода и ангиогенез, увеличивая данные процессы. Некоторые гормоны также способны влиять на экспрессию генов в мышечных клетках. Это такие гормоны, как инсулин, гормон роста, которые вместе с кортизолом запускают катаболические реакции в условиях метаболического и энергетического истощения [3].
Стоит напомнить, что мышцы не являются постоянными клетками, а заменяются в течение жизни. Пролиферация необходима для предотвращения апоптоза клеток (регулируемый процесс клеточной гибели) и поддержания массы скелетных мышц. Это осуществляется через динамический баланс между синтезом белков в мышцах и их распадом. Мышечная гипертрофия возникает тогда, когда синтез белков превышает их распад.
Что же наблюдается при гипертрофии и гиперплазии мышечного волокна? При растяжении и сокращении мышц происходит образование факторов роста IGF и MGF, которые могут действовать как паракринно, так и аутокринно. С одной стороны, их действие проявляется в увеличении синтеза сократительных белков мышечных волокон. Основным участником данного механизма является фосфорилированная PKB [5]. Её активация начинается с влияния на мышцу нагрузки, которая приводит к синтезу гена, запускающего путь IGF/PI3K. В ткани имеется несколько изоформ, некоторые из них (IGF-1 и MGF), взаимодействуя с рецепторами приводят к конформационным изменениям. Через фосфорилирование ряда рецепторов и происходит активация PKB, способствующая развитию анаболических реакций [6].
С другой же стороны, происходит усиление пролиферации миосателлитов, их митотическая активность приводит к формированию новых клеток, а также сопровождается слиянием их с имеющимися мышечными волокнами или даёт возможность формировать новые. Миосателлиты расположены между базальной мембраной и сарколеммой. Покоящиеся клетки активируются непосредственно травмированием мышцы и в ответ на это начинают активно делиться и соединяться с частями поврежденного волокна. Под влиянием тяжёлой изнурительной работы происходит также активация данных клеток из-за образования многочисленных микротравм мышечного волокна. Вследствие этого наблюдается явление подобное процессам, происходящим при воспалении. В зону повреждения активно мигрируют нейтрофилы и макрофаги, которые активируют синтез ранее упомянутых факторов роста, регулирующих пролиферацию и дифференцировку миосателлитов. Мышечная гипертрофия отличается от мышечной гиперплазии. При гипертрофии мышц, увеличиваются сократительные элементы, и межклеточный матрикс расширяется для поддержки роста. Гиперплазия приводит к увеличению количества мышечных волокон. Гипертрофия сократительных элементов может происходить путем добавления саркомеров либо последовательно или параллельно.
В отечественной литературе не утихают споры о патогенетических аспектах мышечного роста. Чаще всего гипертрофию скелетных мышц человека рассматривают как их долговременную адаптацию к физическим нагрузкам различной направленности. Но существует понятие о кратковременной гипертрофии скелетных мышц – то есть изменение объема мышцы в результате одной силовой тренировки. Спортсмены, выступающие в соревнованиях по бодибилдингу или бодифитнесу хорошо знают, что объем мышц можно немного увеличить за счет собственной крови и осмотического давления, если использовать специальный метод тренировки – пампинг.
Неоспоримым является факт увеличения объёма мышечных волокон. Это так называемая миофибриллярная гипертрофия, при которой происходит изменение объёма миофибрилл и плотность их укладки. Механизм связан с увеличением количества саркомеров в миофибриллах. Значительная роль при этом отводится активированным клеткам-сателлитам. Миогенные стволовые клетки начинают пролифелировать, а затем сливаются с существующими клетками или взаимодействуют между собой для формирования новых мышечных волокон. Этот механизм актуален при восстановлении травмированных клеток и при спортивной гипертрофии.
Существует множество данных, доказывающих идущий параллельно с этим процесс увеличения объёма несократительной части мышцы – саркоплазматическая гипертрофия. Это тонкие перестройки на биохимическом уровне клетки, а так же увеличение количества митохондрий. Многие авторы считают, что трансформации в саркоплазме повышают выносливость мышц. Ряд исследователей утверждает, что увеличение различных неконтрактильных элементов и жидкости действительно может привести к приросту мышечной массы, но без сопутствующего увеличения силы. Саркоплазматическая гипертрофия достигается специальными тренировками и часто описывается как нефункциональная. Однако ряд специалистов предполагают, что отек мышечных волокон вызывает увеличение синтеза белка и таким образом способствует росту сократительной ткани.
Эти процессы редко бывают сбалансированными и зависят от характера и интенсивности нагрузки. В скелетных мышцах при этом синтез мышечных белков преобладает над их распадом. Причиной такого метаболизма сторонники гипотезы ацидоза считают накопление молочной кислоты. С точки зрения другой теории – временная гипоксия запускает реперфузию мышц и активирует деление клеток-сателлитов. Последнее время широкое распространение получила гипотеза механического повреждения мышечных волокон. Микроразрывы сократительных белков и повреждения саркоплазмы сопровождается увеличением концентрации ионов кальция, что и стимулирует пролиферацию сателлитов.
Из этого следует, что механизмы мышечной гипертрофии известны и неоспоримы. Очень дискутабельным остается вопрос о наличии процесса гиперплазии мышц. Большинство авторов сходится во мнении, что увеличение количества мышечных волокон у человека не доказано, но при этом описывается возможность получения гиперплазии мышц в экспериментальных условиях у животных (млекопитающих и птиц). Некоторые исследователи допускают частичное увеличения числа волокон. На основании проведенного мета-анализа экспериментальных работ отмечено, что количество мышечных элементов увеличилось в экспериментах на птицах значительнее, чем при использовании в качестве подопытных млекопитающих. Примечательно также, что эффект гиперплазии наблюдался там, где использовались постоянные растяжения, а не упражнения, сочетающие его с расслаблением. Ряд исследователей (Kraemer, William J. и MacDougall J.) утверждают, что этот механизм может осуществляться под влиянием силовых тренировок. Однако доказательств увеличения мышечных волокон у людей недостаточно. Длительных исследований (более года) добровольцев и спортсменов не проводилось. Высказывается мнение, что это слишком короткий период для этого процесса. Гиперплазия подтверждается в биопсийном материале, а погрешность этого метода составляет около 10 %, что делает результат очень сомнительным.
Общее число волокон предопределяется генетически и практически не меняется в течение жизни без применения специальных стимуляторов. Российские ученые подтверждают, что вклад гиперплазии в процесс увеличения объема мышц составляет не более 5 % и, как правило, потенцирован использованием анаболических стероидов. Также гиперплазию могут вызывать блокаторы миостатина. Гормон роста при этом не вызывает гиперплазии.
Таким образом, при мышечной работе происходит множество процессов на разных уровнях. Начиная с изменений интенсивности обменных процессов и заканчивая изменениями механизмов нервной и гуморальной регуляции. Реорганизация мышц, лежащая в основе этих процессов, приводит к изменению многочисленных характеристик деятельности спортсменов.
Проанализировав все данные и изучив все возможные гипотезы, становится очевидным, что в увеличении мышечных волокон играют некую роль всё-таки два процесса. Первый – гипертрофия с ёе подвидами для сократительной и несократительной части мышцы (миофибриллярная и саркоплазматическая), которая, по мнению многих исследователей, занимает основополагающую роль. И второй это гиперплазия с её минимальным, но существенным вкладом.



