Фотосинтез
Типы питания

Фотосинтез

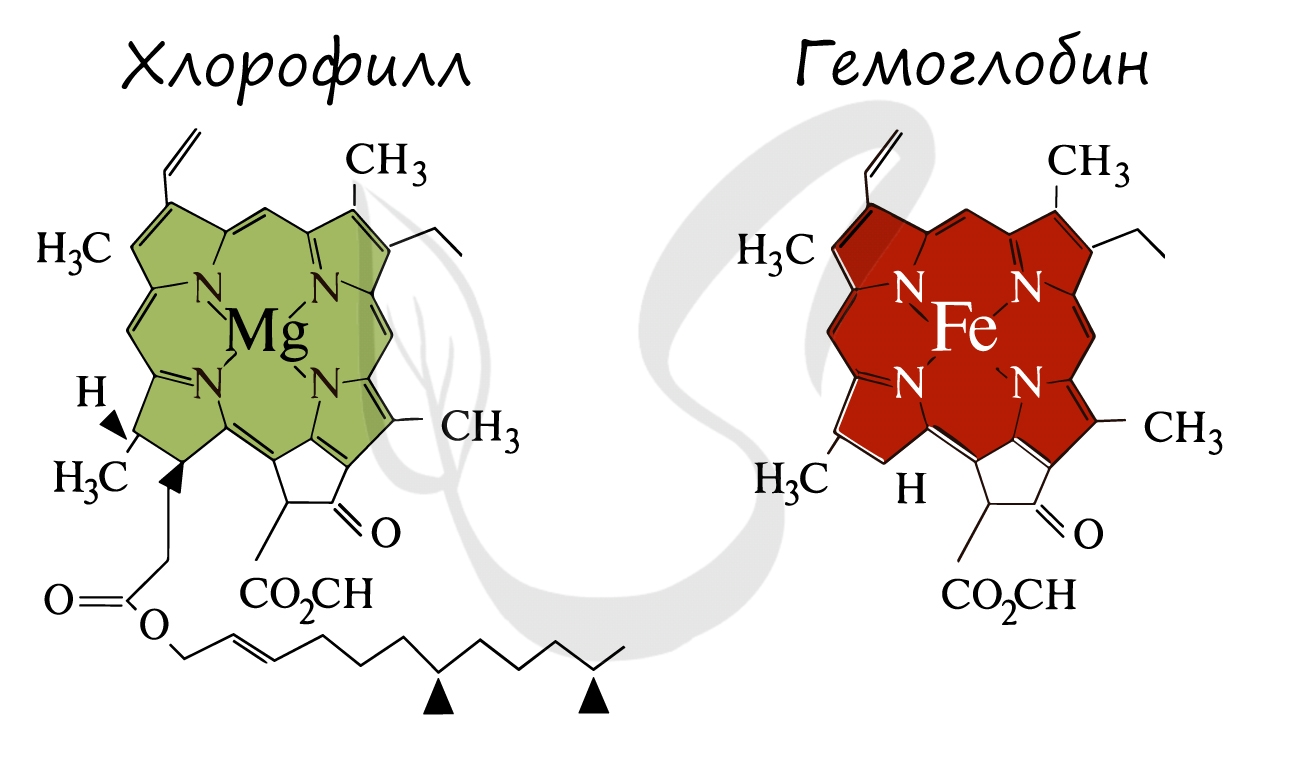
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: «Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического»
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
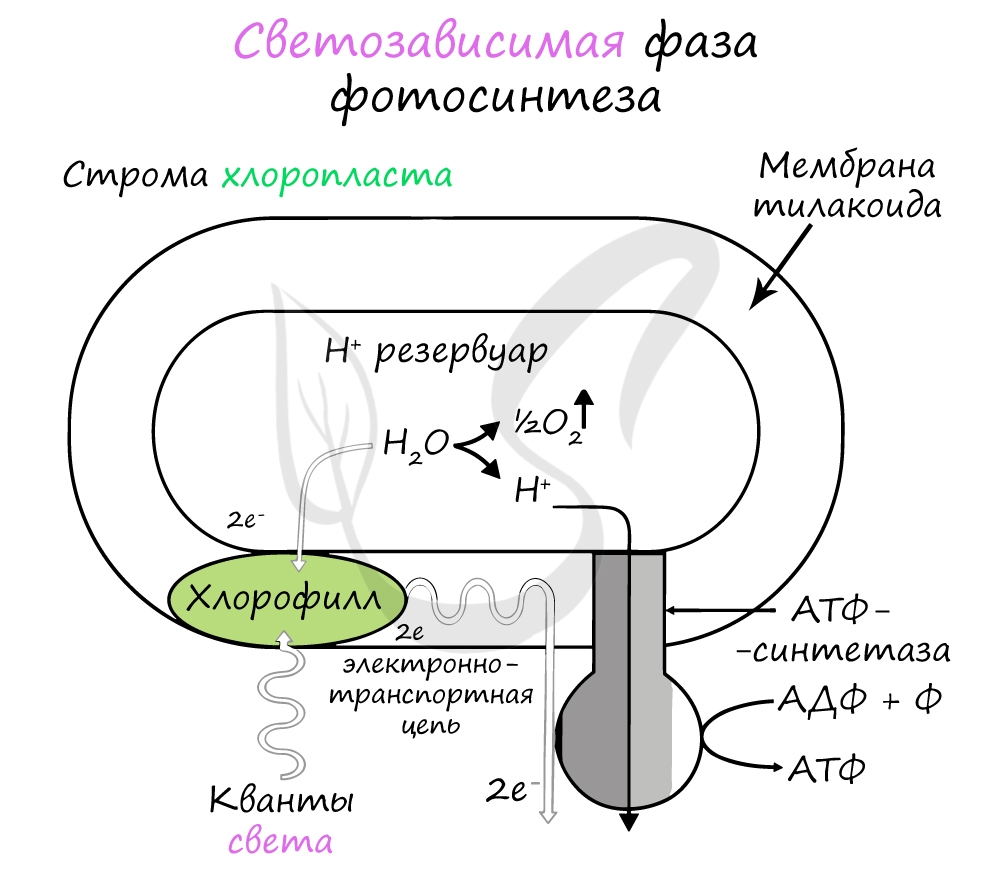
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
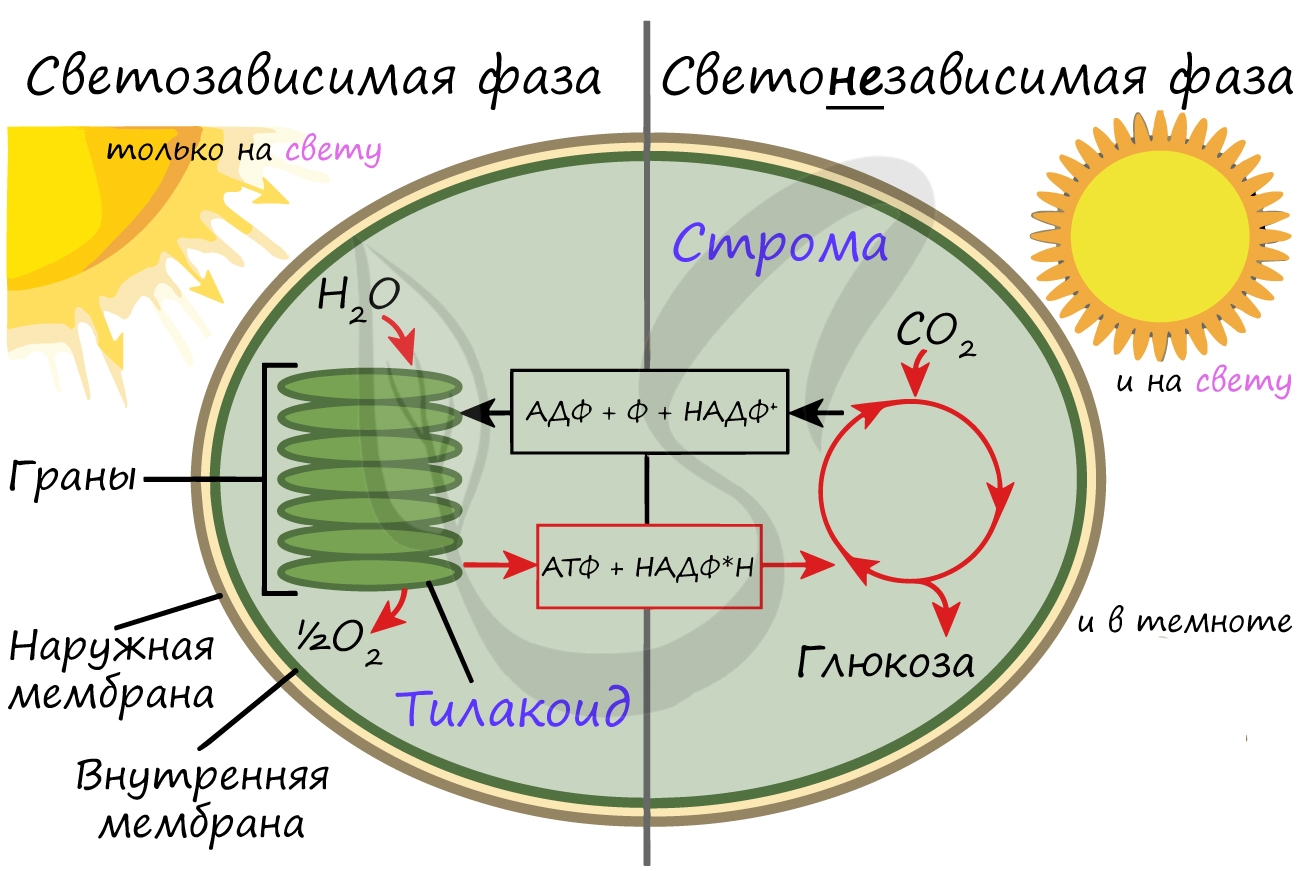
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.

Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
Значение хемосинтеза
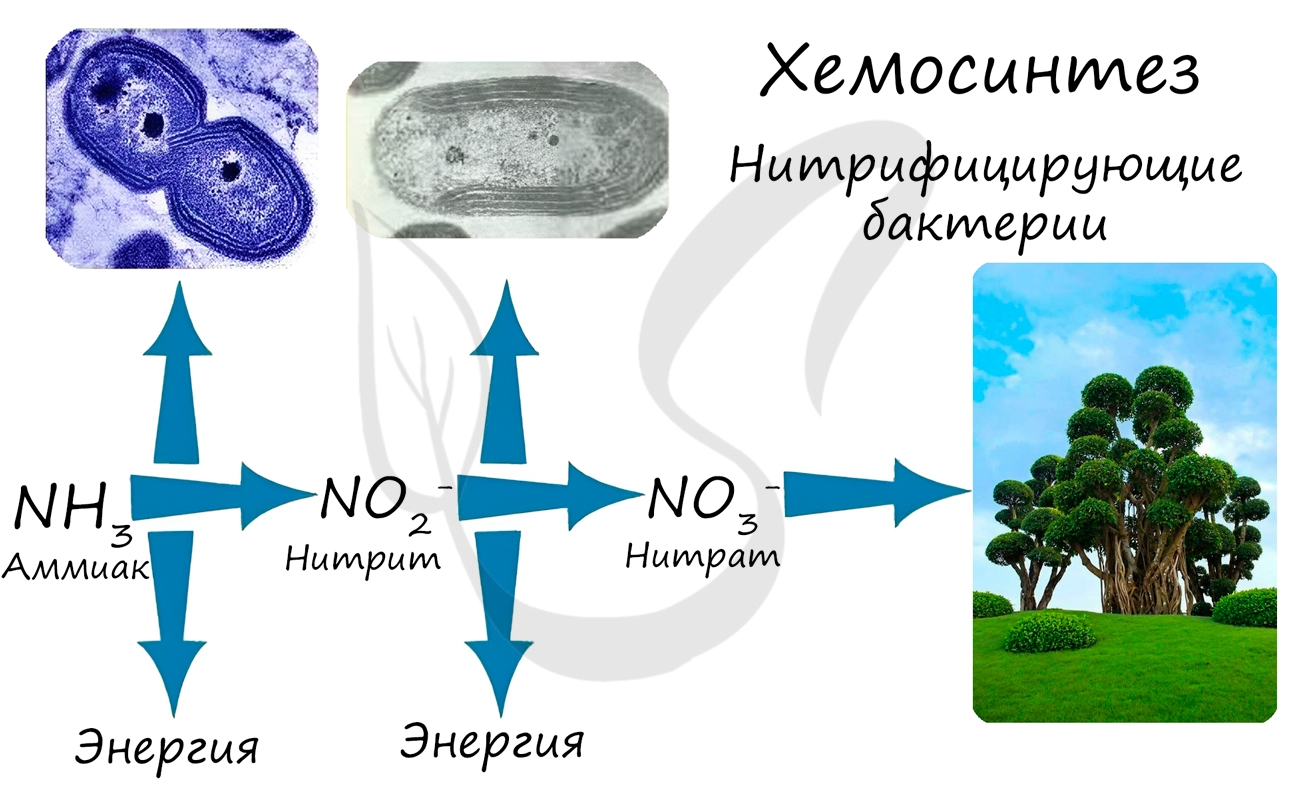
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Лекция № 12. Фотосинтез. Хемосинтез
Фотосинтез
Фотосинтез — синтез органических веществ из углекислого газа и воды с обязательным использованием энергии света:
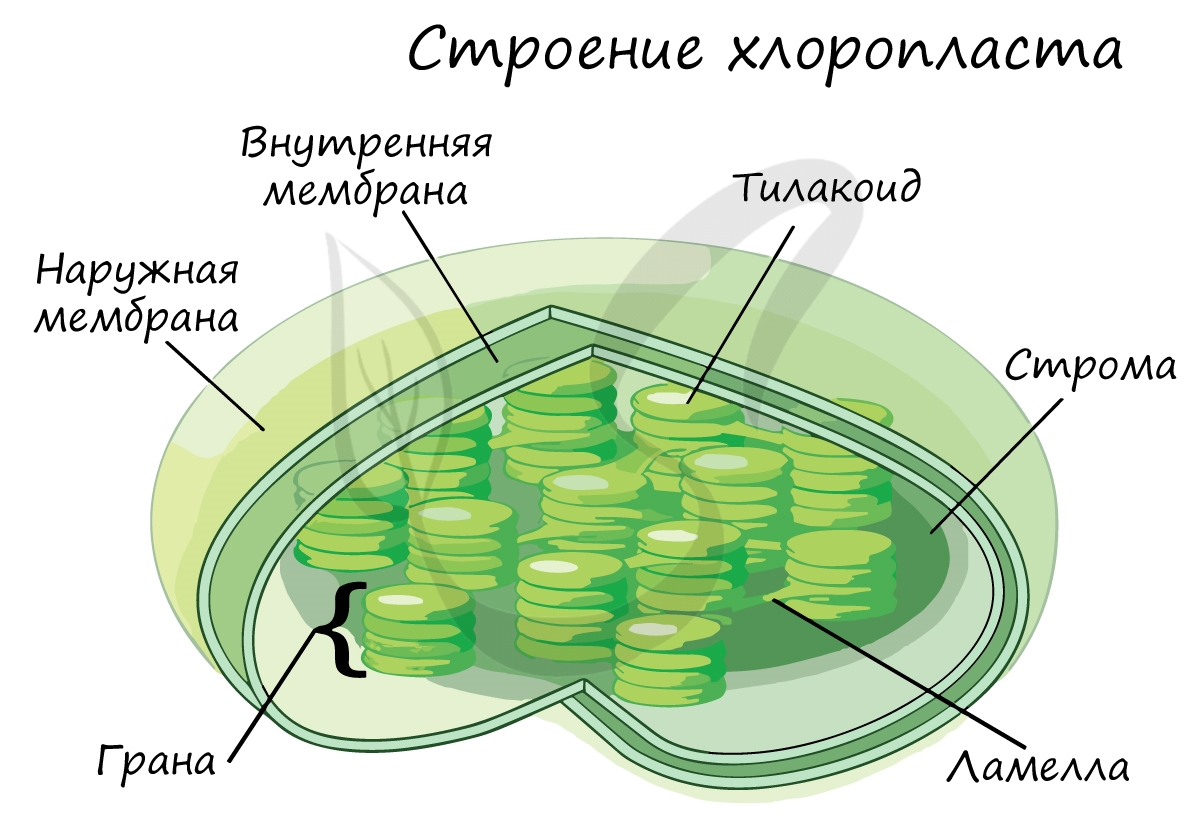
У высших растений органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты (строение хлоропластов — лекция №7). В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру, является гидрофильной и поэтому лежит на той поверхности мембраны, которая обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого удерживает молекулу хлорофилла в мембране.

Хлорофиллы поглощают красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы. У растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
Радикалы •ОН объединяются, образуя воду и свободный кислород:
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет Н + заряжается положительно, с другой за счет электронов — отрицательно. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет на восстановление специфического переносчика НАДФ + (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н + + 2е — + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.

1 — строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является пятиуглеродный сахар рибулозобифосфат (РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется неустойчивое шестиуглеродное соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина»:
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез

Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая фаза». Характерные особенности С3-фотосинтеза: 1) акцептором углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется шестиуглеродное соединение, которое распадается на две ФГК. ФГК восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание

Фотодыхание:
1 — хлоропласт; 2 — пероксисома; 3 — митохондрия.
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать. Он поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
 |  Строение С4-растений: С4-фотосинтез: Значение фотосинтезаКупить проверочные работы
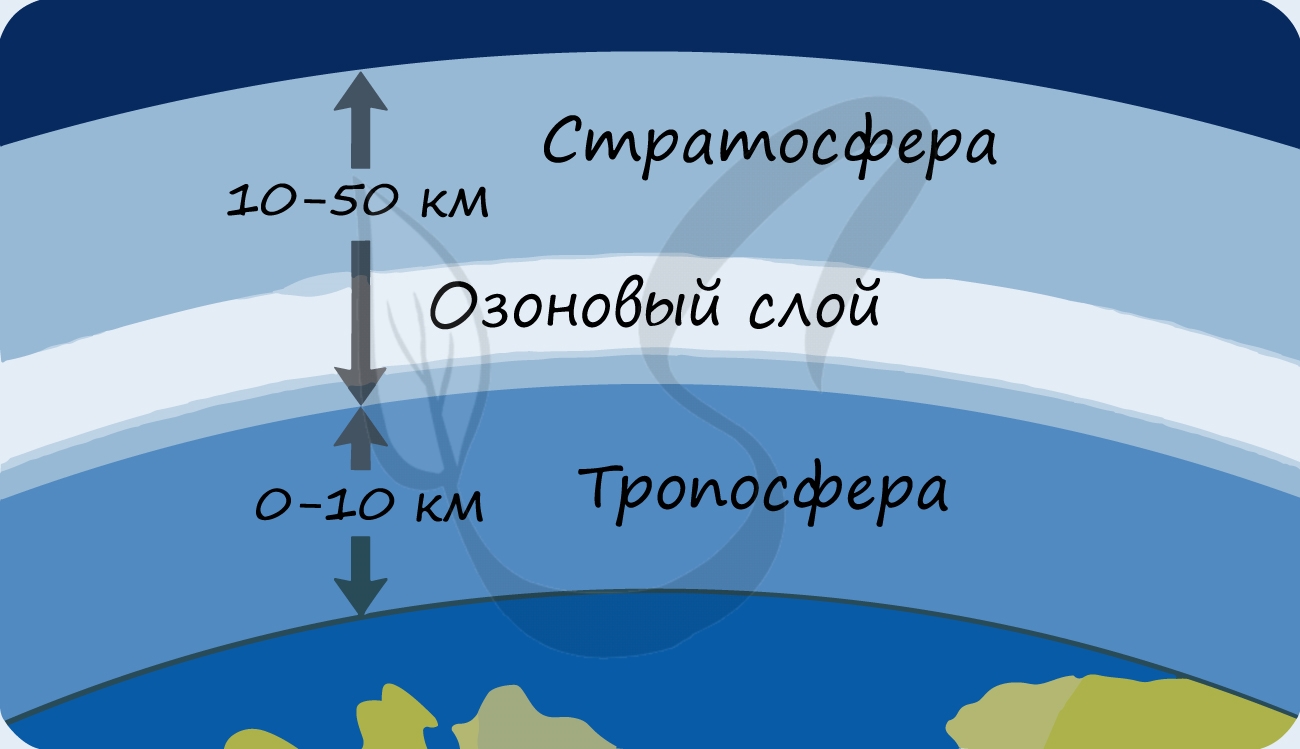
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации. При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м 2 поверхности в час. ХемосинтезСинтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий. Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3). Железобактерии превращают закисное железо в окисное (Fe 2+ → Fe 3+ ). Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4). В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза. Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др. Перейти к лекции №11 «Понятие об обмене веществ. Биосинтез белков» Перейти к лекции №13 «Способы деления эукариотических клеток: митоз, мейоз, амитоз» Смотреть оглавление (лекции №1-25) Фиксация углерода
Хотя углерод в основном фиксируется через 6 автотрофных путей, существуют и неавтотрофные пути. На втором этапе, называемом циклом Кальвина, осуществляется фактическая фиксация углекислого газа. Этот процесс потребляет АТФ и НАДФН. Цикл Кальвина у растений объясняет преобладание фиксации углерода на суше. У водорослей и цианобактерий это объясняет преобладание фиксации углерода в океанах. Цикл Кальвина превращает углекислый газ в сахар в виде триозофосфата (TP), который представляет собой глицеральдегид-3-фосфат (GAP) вместе с дигидроксиацетонфосфатом (DHAP): 3 CO 2 + 6 НАДФ + 6 H + + 9 АТФ + 5 H 2 O → TP + 6 НАДФ + + 9 АДФ + 8 P i Эволюционные соображенияМеханизмы концентрирования CO 2Механизмы концентрирования CO 2 используют фермент карбоангидразу (CA), который катализирует как дегидратацию бикарбоната до CO 2, так и гидратацию CO 2 до бикарбоната. CAM заводыC 4 растенияC 3 растенияБактерии и цианобактерииОбратный цикл КребсаЭтот путь является циклическим из-за регенерации оксалоацетата. [15] Восстановительный путь ацетил-КоАОдна ветвь этого пути, метильная ветвь, похожа, но не гомологична для бактерий и архей. В этой ветви происходит восстановление CO 2 до метильного остатка, связанного с кофактором. Промежуточные продукты представляют собой формиат для бактерий и формилметанофуран для архей, а также носители, тетрагидрофолат и тетрагидроптерины, соответственно, у бактерий и архей, различны, например, ферменты, образующие метильную группу, связанную с кофактором. [14] В противном случае карбонильная ветвь гомологична между двумя доменами и состоит из восстановления другой молекулы CO 2 до карбонильного остатка, связанного с ферментом, катализируемого CO-дегидрогеназой / ацетил-CoA-синтазой. Этот ключевой фермент также является катализатором образования ацетил-КоА, исходя из продуктов предыдущих реакций, метильных и карбонильных остатков. [30] [31] Этот путь фиксации углерода требует только одной молекулы АТФ для производства одной молекулы пирувата, что делает этот процесс одним из основных для хемолитоавтотрофов, ограниченных по энергии и живущих в анаэробных условиях. [14] 3-гидроксипропионатный велосипед3-гидроксипропионатный велосипед состоит из двух циклов, и название этого способа происходит от 3-гидроксипропионата, который соответствует его промежуточной характеристике. Во втором цикле гликоксилат представляет собой примерно одну молекулу пропионил-КоА, образующую метиламалонил-КоА. Это, в свою очередь, затем превращается посредством серии реакций в цитрамалил-КоА. Цитрамалил-КоА расщепляется на пируват и ацетил-КоА благодаря ферменту ММС-лиазе. В этот момент пируват высвобождается, а ацетил-КоА повторно используется и снова карбоксилируется на малонил-коа, таким образом восстанавливая цикл. [35] Это очень дорогой способ: 7 молекул АТФ используются для синтеза нового пирувата и 3 АТФ для фосфаттриозы. [34] Важной характеристикой этого цикла является то, что он обеспечивает совместную ассимиляцию множества соединений, что делает его пригодным для миксотрофных организмов. [34] Два других цикла, связанных с 3-гидроксипропионатным цикломБИОЛОГИЧЕСКИЙ ОТДЕЛ ЦЕНТРА ПЕДАГОГИЧЕСКОГО МАСТЕРСТВА
Фотосинтез. Темновая фазаНа рисунке 3 схематично изображены основные участники темновой фазы фотосинтеза, подверженные регуляции с помощью тиоредоксиновой системы, хотя, несомненно, регуляция гораздо более обширная. На свету восстановленный благодаря работе ЭТЦ хлоропласта пул тиоредоксинов активирует работу ферментов, отвечающих за путь фиксации CO2 и синтеза глюкозы, в то время как активность ферментов путей анаэробного дыхания – окисления глюкозы (гликолиз и пентозофосфатный путь), будут ингибирована. В темноте ситуация противоположная: тиоредоксиновый пул весь окислен, восстановить его нечем, ферменты пути фиксации углерода не активируются, зато протекают пути окисления синтезированной ранее на свету глюкозы. Помимо тиоредоксиновой регуляции, как уже было сказано, наблюдается регуляция с помощью pH. Те же ферменты, что активируются днем тиоредоксинами, увеличивают свою активность и благодаря повышенным дневным значениям pH стромы хлоропласта (pH=8.0). Рис 3. Светозависимая регуляция компонентов темновой фазы фотосинтеза. Цикл Кальвина состоит из трех стадий: 1) карбоксилирование; 2) восстановление; 3) регенерация. Стадия карбоксилирования состоит из одной реакции, катализируемой рибулозобисфосфат карбоксилазой-оксигеназой или RuBisCO. Пожалуй, RuBisCO – самый распространенный фермент на Земле и при этом очень древний. Содержание RuBisCO может составлять порядка 50% массы растворимых белков зеленых листьев, при этом концентрация фермента на порядки выше концентрации субстрата – СО2, что абсолютно нехарактерно для ферментативных реакций. У растений, большинства водорослей и фотосинтетических бактерий RuBisCO состоит из 8 больших субъединиц (L, 55 kDa), кодируемых пластидным геномом и 8 малых субъединиц (S, 13 kDa), кодируемых ядерным геномом. Большие субъединицы являются каталитическими, малые – регуляторными, и они не являются необходимыми для самой реакции карбоксилирования. L субъединицы организованы в димеры, при этом активные сайты карбоксилирования образованы аминокислотами обеих частей димера, таким образом, RuBisCO с формулой L8S8 содержит 8 активных сайтов (бывают и другие типы RuBisCO). Помимо описанного для активации RuBisCO необходимо взаимодействие с активазой RuBisCO – АТР-зависимым ферментом (т.е. тоже работает только на свету!). В темноте в активном сайте RuBisCO, не образовавшем карбамата, прочно удерживается RuBP, который блокирует работу фермента. Активаза RuBisCO высвобождает RuBP из активного сайта, делая его доступным для образования карбамата. Более подробный механизм реакций, происходящих в активном сайте RuBisCO, можно посмотреть здесь: https://www.ebi.ac.uk/thornton-srv/m-csa/entry/907/. На следующей стадии, стадии восстановления, происходят процессы, обратные реакциям гликолиза (рисунок 7): фосфорилирование 3-фосфоглицерата фосфоглицераткиназой до 1,3-дифосфоглицерата и затратой одной молекулы ATP, и дальнейшее восстановление 1,3-дифосфоглицерата ферментом глицеральдегидфосфатдегидрогеназой до глицеральдегид-3-фосфата с использованием в качестве донора электронов NADPH+H + и выделением свободного фосфата. Затем из глицеральдегид-3-фосфата в равновесной реакции, катализируемой триозофосфатизомеразой образуется дигидроксиацетонфосфат. Использование в реакциях стадии восстановления ATP и NADPH+H + делает стадию сильно зависимой от энергии, а значит – и от солнечного света. Образованные восстановленные триозы далее вступают в реакции стадии регенерации. На этой стадии происходят три основных типа реакций – альдолазная, транскетолазная и фосфатазная. Основные процессы схематично изображены на рисунке 8. Сначала в реакции, катализируемой ферментом альдолазой из глицеральдегид-3-фосфата и дигидроксиацетонфосфата образуется фруктозо-1,6-бисфосфат. Альдолазная реакция обратима, в ее механизме в активном центре фермента происходит образование протонированного Шиффового основания между С2 кетозы (дигидроксиацетонфосфата) и ε-аминогруппой остатка лизина, в ходе дальнейшей нуклеофильной реакции с С1 карбонильной группы альдозы (глицеральдегид-3-фосфата) образуется фруктозо-1,6-бисфосфат. Далее фруктозо-1,6-бисфосфат гидролизуется до фруктозо-6-фосфата. Эту реакцию катализирует фруктозо-1,6-бисфосфатаза, и данная реакция необратима. Необратимость реакции позволяет сделать ее регуляторной для всего цикла. Далее фруктозо-6-фосфат вступает с глицеральдегид-3 фосфатом в транскетолазную реакцию, в ходе которой на глицеральдегид-3-фосфат переносятся С1-С2 с фруктозо-6-фосфата. В данной реакции участвует кофермент тиаминпирофосфат (TPP), являющийся простетической группой транскетолазы. ТРР, связываясь углеродом тиазольного кольца с С2 атомом кетозы, вызывает расщепление ковалентной связи между С2 и С3 на кетозе и образование карбаниона с отрицательным зарядом на С2, к которому присоединяется С-атом альдегидной группы альдозы (глицеральдегид-3-фосфат), имеющий частичный положительный заряд. В ходе реакции образуются четырехуглеродный сахар эритрозо-4-фосфат и пентоза – ксилулозо-5-фосфат. Образованный эритрозо-4-фосфат вступает в альдолазную реакцию с дигидроксиацетонфосфатом с образованием седогептулозо-1,7-бисфосфата, который затем гидролизуется до седогептулозо-7-фосфата. Реакция во многом похожа на гидролиз фруктозо-1,6-бисфосфата и так же является необратимой и регуляторной. В следующей транскетолазной реакции седогептулозо-7-фосфат взаимодействует с глицеральдегид-3-фросфатом с образованием двух пентоз – рибозо-5-фосфата и ксилулозо-5-фосфата. Итого, получены 3 различные пентозы. В последующих реакциях (рисунок 9) происходит эпимеризация ксилулозо-5-фосфата в рибулозо-5-фосфат под действием фермента рибулозофосфатэпимеразы и изомеризация рибозо-5-фосфата в рибулозо-5-фосфат под действием фермента рибозофосфатизомеразы. Полученные рибулозо-5-фосфаты фосфорилируются по С1 рибулозофосфаткиназой с образованием рибулозо-1,5-бисфосфата (RuBP). Данная реакция необратима. Таким образом, в ходе цикла Кальвина вновь образовался RuBP, который снова может вступать в реакцию карбоксилирования. Помимо этого был зафиксирован СО2. Если сделать пересчет на три зафиксированных СО2, то есть – на одну синтезированную триозу, то суммарное уравнение ОПФП выглядит следующим образом: 3CO2 + 6NADPH + 6H + + 9ATP → глицеральдегид-3-фосфат + 6NADP + + 9ADP + 3H2O + 8Pi Кроме восстановительного пентозофосфатного пути существует еще и окислительный. Большинство ферментов и реакций этих путей общие и многие реакции являются обратимыми, ОПФП, как и ВПФП проходит в строме хлоропласта. Очевидно, что для нормального функционирования обоих путей и во избежание бессмысленной траты АТP необходимо регулировать активность ферментов, которая осуществляется с помощью тиоредоксиновой системы и изменения рН стромы хлоропласта (рис 10). Помимо карбоксилазной активности RuBisCO проявляет также и оксигеназную (рисунок 11). В ходе этой реакции в активный центр RuBisCO вместо СО2 приходит О2 и образуются 2-фосфогликолат и только один 3-фосфоглицерат. Даже небольшие концентрации 2-фосфогликолата опасны для растения, вещество оказывает сильное и ингибирующее воздействие на многие системы и реакции. Для устранения последствий карбоксилазной активности RuBisCO в растениях существует целый метаболический путь, называемый фотодыханием, или гликолатный путь (не путать с глиоксилатным!), или С2-фотосинтез. Реакции данного пути требуют консолидированной работы сразу трех органелл – хлоропласта, пероксисомы и митохондрии (рисунок 12, из учебника Страсбургера «Физиология растений»). Образованный в строме хлоропласта 2-фосфогликолат гидролизуется фосфогликолатфосфатазой и продукт, гликолат, выносится из хлоропласта через интегрированныйво внутреннюю мембрану транспортер и попадает в пероксисому через образованные поринами пор. В пероксисоме гликолат окисляется до глиоксилата гликолатоксидазой. Данная реакция идет в присутствии кислорода, на который через флафинмононуклеотид гликолатоксидазы передаются электроны и образуется H2O2. Н2О2 разлагается каталазой до воды и кислорода, именно поэтому данную реакцию необходимо проводить в пероксисоме. Далее ферментом глутамат-глиоксилатаминотрансферазой осуществляется переаминирование – перенос аминогруппы с глутамата на глиоксилат с образованием 2-оксоглутарата и глицина. В качестве донора аминогруппы может использоваться также аланин, помимо этого реакцию осуществляет и серин-глиоксилатаминотрансфераза (см. далее). Глицин затем поступает через аминокислотный транслокатор в митохондрию, где окисляется мультиферментным комплексом, сходным с пируватдегидрогеназным комплексом цикла Кребса, полное название которого – глициндекарбоксилазосерингидроксиметилтрансферазный комплекс (рисунок 13). В ходе реакций, катализируемых данным комплексом глицин, связываясь с пиридоксалем, образует основание Шиффа (B) и декарбоксилируется, затем остаток глицина передается на липоевую кислоту (C) и дезаминируется, окисляясь до формильного остатка, который связывается с тетрагидрофолатом (D, 1, 2). Восстановленная при этом дигидролипоевая кислота окисляется флафинадениндинуклеотидом (FAD) снова до липоевой кислоты, при этом FADH2 восстанавливает NADP + (D-E). Формильный остаток с тетрагидрофолата передается на вторую молекулу глицина, образуя серин (А-В). Аммоний, образованный в ходе дыхания, также переносится в хлоропласт, где с использованием ATP ферментом глутаминсинтетазой осуществляется аминирование глутамита до глутамина, а затем глутаматоксоглутаратаминотрансфераза (ГОГАТ) из глутамина и 2-оксоглутарата при использовании двух восстановленных ферредоксинов (то есть реакция фиксации аммония в листьях тоже светозависима!) в качестве доноров электронов катализирует образование двух глутаматов.
Затраты на соответствующую реакцию на 1 моль газа Затраты (моль) Карбоксилазная Оксигеназная Состояние, когда весь фиксируемый CO2 расходуется из-за оксигеназной активности, получило название углекислотной компенсации. Нетрудно догадаться из стехиометрии ВПФП и фотодыхания, что такая ситуация будет наблюдаться при соотношении карбоксилазной и оксигеназной реакций = 1:2. Рассмотренные процессы происходят в растениях с так называемым С3 фотосинтезом, по количеству углеродов в первом образованном после фиксации СО2 веществе – 3-фосфоглицерате. Но у растений существуют и другие типы фотосинтеза, их много, если учесть переходные формы. Далее мы рассмотрим некоторые основные. Концентрация кислорода в современной атмосфере равна 21%, СО2 – порядка 0,035-0,04%. Известно, что с повышением температуры растворимость газов уменьшается, а так как СО2 всего лишь около 350 ppm, в водном растворе СО2 становится очень мало по сравнению с содержанием О2, падает соотношение CO2/O2, что способствует значительному возрастанию оксигеназной активности. Растения научились бороться с этим и выживать в засушливых условиях с повышенной температурой и инсоляцией. Ключевая реакция здесь – первичная фиксация углекислого газа ферментом фосфоенолпируваткарбоксилазой (ФЕП-карбоксилаза, PEPCase). В одном случае CO2 в форме карбоксильной группы низкомолекулярных органических веществ (малат, оксалоацетат, аспартат) перемещается из клетки первичной фиксации (клетка хлоренхимы мезофилла) в клетку обкладки пучка (рис. 14), где происходит декарбоксилирование и образование СО2. Тут СО2 уже фиксируется с помощью RuBisCO. Данный механизм позволяет создать в области активности RuBisCO условия с повышенной концентрацией СО2, благодаря чему можно если не убрать, то минимизировать оксигеназную реакцию. Схематичный процесс, который называется в честь первооткрывателей циклом Хэтча-Слэка-Карпилова (хотя последнего все забывают), представлен на рисунке 14. Такой фотосинтез называется С4 из-за того, что первоначально при первичной фиксации образуется четырехуглеродное соединение – оксалоацетат. Листья растений с С4 типом фотосинтеза имеют своеобразную анатомию (рисунок 15), называемую кранц-анатомией (нем. Kranz – венок). Кранц-анатомия и С4 фотосинтез эволюционно возникала независимо во многих таксонах цветковых растений и имеет различное строение в зависимости от типа С4 фотосинтеза и видовой принадлежности самого растения, более того, даже помимо переходных случаев, вроде С3-С4 фотосинтеза, есть данные, что у типичных С3 растений вполне может проходить и С4 фотосинтез. ФЕП-карбоксилаза катализирует образование оксалоацетата из иона бикарбоната и фосфоенолпирувата. На первой стадии реакции происходит нуклеофильная атака бикарбонатом фосфата фосфоенолпирувата, образуются два промежуточных продукта – фосфокарбонат и енольная форма пирувата. На второй, необратимой стадии, происходит разрыв эфирной связи в фосфокарбонате, выходит из реакции Pi, а CO2 реагирует с енольной формой пирувата с образованием оксалоацетата. У С4 растений ФЕП-карбоксилаза активируется фосфорилированием осуществляемым ферментом киназой ФЕП-карбоксилазы, при этом сама киназа активируется светом, в темноте ФЕП карбоксилаза инактивируется фосфорилазой, гидролизующей присоединенный на свету фосфат. Поэтому данная стадия у С4 растений идет только на свету. У ФЕП-карбоксилазной реакции два больших преимущества: (1) отсутствие оксигеназной активности и (2) использование бикарбоната в качестве субстрата. Образование HCO3— из СО2 (помимо самопроизвольного) катализируется карбоангидразой. Данный факт позволяет накопить большое количество углерода в форме HCO3-, ведь в равновесии с 8 мкмоль СО2 при 25°С и рН=8 находится 400 мкмоль HCO3-, таким образом, минимизируется проблема малого количества растворенного СО2 при высокой температуре. Есть и большое количество других переносчиков, обменивающих аминокислоты, фосфаты, нуклеотиды, ионы и проч. С4 тип фотосинтеза происходит в основном у растений, обитающих в климате с повышенной температурой и отчасти с недостатком воды (рис 23). Если брать в рассмотрение только протекающие в процессе фиксации углерода реакции, то можно прийти к выводу, что по сравнению с C3 фотосинтезом С4 довольно расточителен. Действительно, в условиях умеренного климата с относительно стабильным и достаточным увлажнением растения с С3 фотосинтезом имеют преимущество перед С4 растениями, опережая их в скорости накопления биомассы. Но как только растения оказываются в жарком климате, энергетические затраты на дополнительные реакции у С4 растений м перекрываются большим выигрышем в уровне фиксации СО2 и уменьшении оксигеназной активности RuBisCO. Растения, произрастающие в засушливых с долговременной нехваткой влаги или даже полным отсутствием воды в течение продолжительного времени, развили третий тип фотосинтеза. CAM-фотосинтез (от англ. Crassulacean acid metabolism – метаболизм по типу толстянковых) довольно широко распространен среди высших растений, хоть и был впервые описан на семействе Crassulaceae. САМ фотосинтез, точно так же, как и С4, возникал у разных групп растений неоднократно и независимо друг от друга. Растения с САМ фотосинтезом встречаются и во многих группах – представителей можно встретить среди плауновидных, папоротникообразных, гнетовых, голосеменных и покрытосеменных растений. Точно так же есть растения с переходными типами фотосинтеза, существует много примеров изменения типа фотосинтеза с С3 на САМ у одного растения в зависимости от внешних условий и/или стадии онтогенеза и даже от рассматриваемого органа. САМ фотосинтез в основном выполняет роль физиологической адаптации к недостатку воды, потому встречается не только у пустынных растений, но так же у эпифитов и галофитов. Помимо этого, САМ фотосинтез имеет распространение и среди водных растений (например, Isoetes howellii или Crassula aquatica). Недостатка воды в данном случае растение не испытывает, но сталкивается с недостатком СО2 в силу его низкой концентрации и малой доступности, особенно в кислых водоемах, где равновесие сдвинуто в сторону образования СО2 из растворенного бикарбоната. Такие растения фиксируют СО2 ночью, когда не так высока конкуренция за СО2 с другими фотосинтетиками, населяющими водоем, а также когда эти фотосинтетики не образуют кислород. САМ фотосинтез с точки зрения биохимии процессов очень похож на С4, основная его особенность в том, что происходит разделение первичной и вторичной фиксации СО2 не в пространстве, а во времени. В данном случае первичная фиксация с помощью ФЕП-карбоксилазы происходит ночью, в вечерние и ранние утренние часы – то есть в то время, когда у растений открыты устьица, а активность RuBisCO наблюдается днем, когда устьица закрыты, чтобы максимально уменьшить испарение воды. В отличие от С4 типа фотосинтеза, в данном случае ФЕП-карбоксилаза работает не на свету, а ночью и светом наоборот ингибируется. Работа данного варианта фермента контролируется циркадными ритмами на уровне транскрипции киназы ФЕП-карбоксилазы – транскрипты этого фермента накапливаются именно в темное время суток.  Как 16-летний парень, я бы хотел поделиться своим восторгом  Если вы ищете игру, которая бросит вас в самое сердце |









