Как называется бластная форма в эритроцитарном диффероне
ГЕМОПОЭЗ = (или, КРОВЕТВОРЕНИЕ), это образование форменных элементов крови из ПОЛИПОТЕНТНОЙ СТВОЛОВОЙ КРОВЕТВОРНОЙ КЛЕТКИ (ПСКК).
Она находится в красном костном мозге.
Из нее развиваются ВСЕ форменные элементы (эритроциты, лейкоциты и тромбоциты).
Группа клеток от самой НЕзрелой (т.е. НЕдифференцированной ) до самой зрелой (напр. от ПСКК до эритроцита) назыв. клеточный ДИФФЕРОН..
Гемопоэз после рождения назыв. ПОСТНАТАЛЬНЫЙ гемопоэз.
Он происх. в красном костном мозге (эритроцитопоэз, моноцитопоэз, тромбоцитопоэз),
а развитие Т-лимфоцитов происх. в ТИМУСЕ (Т-лимфоцитопоэз)
Развитие эритроцитов происх. в ЭРИТРОБЛАСТИЧЕСКОМ ОСТРОВКЕ. Островок состоит из МАКРОФАГА красного костного мозга и ЭРИТРОБЛАСТОВ. У макрофага есть длинные отростки цитоплазмы. К ним прикрепляются эритробласты. Макрофаг передает им белок ФЕРРИТИН, который они используют для синтеза гемоглобина.
Развитие тромбоцитов назыв. ТРОМБОЦИТОПОЭЗ.
Дифферон тромбоцитопоэза:
ПСКК – КОЕ-ГЭММ – мегакариоцит – тромбоцит.
Развитие тромбоцитов происх. в ОСТРОВКЕ ТРОМБОЦИТОПОЭЗА.
Островок состоит из синусоидного капилляра и мегакариоцита.
Мегакариоцит – самая БОЛЬШАЯ кроветворная клетка. Она нах. снаружи от синусоидного капилляра и просовывает отростки цитоплазмы в просвет капилляра. От отростков отщепляются фрагменты и они становятся тромбоцитами.
Значит, тромбоциты – это фрагменты цитоплазмы мегакариоцита.
Гемопоэз у зародыша назыв. ПРЕНАТАЛЬНЫЙ гемопоэз.
Самым ранним (первым) кроветворным органом является ЖЕЛТОЧНЫЙ МЕШОК.
В МЕЗОДЕРМЕ желточного мешка развиваются первые кровеносные сосуды и первичные эритроциты, которые назыв. МЕГАЛОБЛАСТЫ. Это назыв. мезобластическая стадия гемопоэза.
Гистология. Полный курс за 3 дня.
Тема 11. КРОВЕТВОРЕНИЕ.
Кроветворение (гемоцитопоэз) – процесс образования форменных элементов крови.
Различают два вида кроветворения:
В свою очередь миелоидное кроветворение подразделяется на:
Лимфоидное кроветворение подразделяется на:
Кроме того, гемопоэз подразделяется на два периода:
Эмбриональный период приводит к образованию крови как ткани и потому представляет собой гистогенез крови. Постэмбриональный гемопоэз представляет процесс физиологической регенерации крови как ткани.
Эмбриональный период гемопоэза.
Он осуществляется в эмбриогенезе поэтапно, сменяя разные органы кроветворения. В соответствии с этим выделяют три этапа:
1. Желточный этап осуществляется в мезенхиме желточного мешка начиная со 2 – 3-й недели эмбриогенеза, с 4-й – снижается и к концу 3-го месяца полностью прекращается.
Вначале в желточном мешке в результате пролиферации мезенхимальных клеток образуются так называемые кровяные островки, представляющие собой очаговые скопления отростчатых клеток.
Наиболее важными моментами желточного этапа являются:
1) образование стволовых клеток крови;
2) образование первичных кровеносных сосудов.
Несколько позже (на 3-й неделе) начинают формироваться сосуды в мезенхиме тела зародыша, однако они являются пустыми щелевидными образованиями. Довольно скоро сосуды желточного мешка соединяются с сосудами тела зародыша, и устанавливается желточный круг кровообращения. Из желточного мешка по этим сосудам стволовые клетки мигрируют в тело зародыша и заселяют закладки будущих кроветворных органов (в первую очередь печень), в которых затем и осуществляется кроветворение.
2. Гепатотимусолиенальный этап) гемопоэза осуществляется вначале в печени, несколько позже в тимусе (вилочковой железе), а затем и в селезенке. В печени происходит (только экстраваскулярно) в основном миелоидное кроветворение начиная с 5-й недели и до конца 5-го месяца, а затем постепенно снижается и к концу эмбриогенеза полностью прекращается. Тимус закладывается на 7 – 8-й неделе, а несколько позже в нем начинается Т-лимфоцитопоэз, который продолжается до конца эмбриогенеза, а затем и в постнатальном периоде до его инволюции (в 25 – 30 лет). Селезенка закладывается на 4-й неделе, с 7 – 8-й недели она заселяется стволовыми клетками, и в ней начинается универсальное кроветворение, т. е. и миело– и лимфопоэз. Особенно активно кроветворение протекает в селезенке с 5-го по 7-й месяцы, а затем миелоидное кроветворение постепенно угнетается, и к концу эмбриогенеза (у человека) оно полностью прекращается.
3. Медуллотимусолимфоидный этап кроветворения. Закладка красного костного мозга начинается со 2-го месяца, кроветворение в нем начинается с 4-го месяца, а с 6-го месяца он является основным органом миелоидного и частично лимфоидного кроветворения, т. е. является универсальным кроветворным органом. В это же время в тимусе, селезенке и в лимфатических узлах осуществляется лимфоидное кроветворение.
В результате последовательной смены органов кроветворения и совершенствования процесса кроветворения формируется кровь как ткань, которая у новорожденных имеет существенные отличия от крови взрослых людей.
Постэмбриональный период кроветворения.
Осуществляется в красном костном мозге и лимфоидных органах (тимусе, селезенке, лимфоузлах, миндалинах, лимфоидных фолликулах).
Сущность процесса кроветворения заключается в пролиферации и поэтапной дифференцировке стволовых клеток в зрелые форменные элементы крови.
В схеме кроветворения представлены два ряда кроветворения:
Каждый вид кроветворения подразделяется на разновидности (или ряды) кроветворения.
1) эритроцитопоэз (или эритроцитарный ряд);
2) гранулоцитопоэз (или грануляцитарный ряд);
3) моноцитопоэз (или моноцитарный ряд);
4) тромбоцитопоэз (или тромбоцитарный ряд).
1) Т-лимфоцитопоэз (или Т-лимфоцитарный ряд;
В процессе поэтапной дифференцировки стволовых клеток в зрелые форменные элементы крови в каждом ряду кроветворения образуются промежуточные типы клеток, которые в схеме кроветворения составляют классы клеток.
Всего в схеме кроветворения различают шесть классов клеток.
I класс – стволовые клетки. По морфологии клетки этого класса соответствуют малому лимфоциту. Эти клетки являются полипотентными, т. е. способны дифференцироваться в любой форменный элемент крови. Направление дифференцировки зависит от содержания форменных элементов в крови, а также от влияния микроокружения стволовых клеток – индуктивных влияний стромальных клеток костного мозга или другого кроветворного органа. Поддержание популяции стволовых клеток осуществляется следующим образом. После митоза стволовой клетки образуются две: одна вступает на путь дифференцировки до форменного элемента крови, а другая принимает морфологию лимфоцита малого размера, остается в костном мозге, является стволовой. Деление стволовых клеток происходит очень редко, их интерфаза составляет 1 – 2 года, при этом 80% стволовых клеток находятся в состоянии покоя и только 20% – в митозе и последующей дифференцировке. Стволовые клетки также получили название колинеобразующие единицы, так как каждая стволовая клетка дает группу (или клон) клеток.
II класс – полустволовые клетки. Эти клетки являются ограниченно полипотентными. Выделяют две группы клеток – предшественницы миелопоэза и лимфопоэза. По морфологии похожи на малый лимфоцит. Каждая из этих клеток дает клон миелоидного или лимфоидного ряда. Деление происходит раз в 3 – 4 недели. Поддержание популяции осуществляется аналогично полипотентным клеткам: одна клетка после митоза вступает в дальнейшую дифференцировку, а вторая остается полустволовой.
III класс – унипотентные клетки. Данный класс клеток является поэтинчувствительными – предшественниками своего ряда кроветворения. По морфологии они также соответствуют малому лимфоциту и способны к дифференцировке только в один форменный элемент крови. Частота деления данных клеток зависит от содержания в крови поэтина – биологически активного вещества, специфического для каждого ряда кроветворения, – эритропоэтина, тромбоцитопоэтина. После митоза клеток данного класса одна клетка вступает в дальнейшую дифференцировку до форменного элемента, а вторая поддерживает популяцию клеток.
Клетки первых трех классов объединяются в класс морфологически не идентифицируемых клеток, так как все они по морфологии напоминают малый лимфоцит, однако способности их к развитию различны.
IV класс – бластные клетки. Клетки этого класса отличаются по морфологии от всех остальных. Они крупные, имеют крупное рыхлое ядро (эухроматин) с 2 – 4 ядрышками, цитоплазма базофильна за счет большого количества свободных рибосом. Эти клетки часто делятся, и все дочерние вступают в дальнейшую дифференцировку. Бласты различных рядов кроветворения можно идентифицировать по цитохимическим свойствам.
V класс – созревающие клетки. Этот класс характерен для своего ряда кроветворения. В этом классе может быть несколько разновидностей переходных клеток от одной (пролимфоцит, промоноцит) до пяти в эритроцитарном ряду. Некоторые созревающие клетки в небольшом количестве могут попадать в периферический кровоток, например ретикулоциты или палочкоядерные лейкоциты.
VI класс – зрелые форменные элементы. К этому классы относятся эритроциты, тромбоциты и сегментоядерные гранулоциты. Моноциты не являются окончательно дифференцированными клетками. Они затем покидают кровеносное русло и дифференцируются в конечный класс – макрофаги. Лимфоциты дифференцируются в конечный класс при встрече с антигенами, при этом они превращаются в бласты и снова делятся.
Совокупность клеток, составляющих линию дифференцировки стволовой клетки в определенный форменный элемент, образует дифферон (или гистогенетический ряд). Например, эритроцитарный дифферон составляют:
1) стволовая клетка (I класс);
2) полустволовая клетка – предшественница миелопоэза (II класс);
3) унипотентная эритропоэтинчувствительная клетка (III класс);
4) эритробласт (IV класс);
5) созревающая клетка – пронормоцит, базофильный нормоцит, полихроматофильный нормоцит, оксифильный нормоцит, ретикулоцит (V класс);
6) эритроцит (VI класс).
В процессе созревания эритроцитов в V классе происходят синтез и накопление гемоглобина, редукция органелл и клеточного ядра. В норме пополнение эритроцитов осуществляется за счет деления и дифференцировки созревающих клеток – пронормоцитов, базофильных и полихроматофильных нормоцитов. Такой тип кроветворения получил название гомопластического. При выраженной кровопотере пополнение эритроцитов осуществляется не только усилением созревающих клеток, но и клеток IV, III, II и даже I класса – происходит гетеропластический тип кроветворения.
Как называется бластная форма в эритроцитарном диффероне
Стадии дифференцировки красных клеток крови. Первой клеткой, которую можно идентифицировать как принадлежащую к группе красных клеток крови, является проэритробласт, показанный как отправная точка на рисунке ниже.
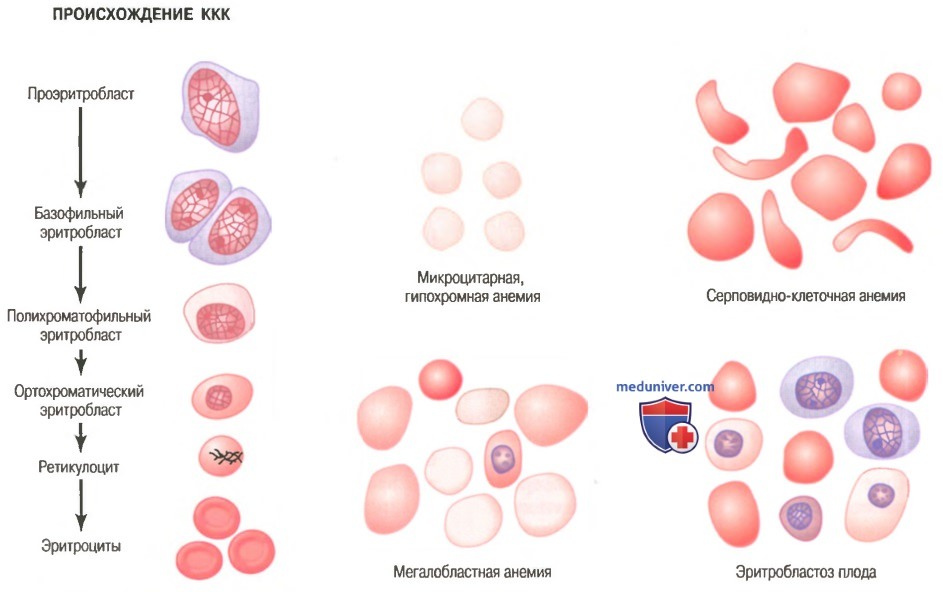 Происхождение нормальных красных клеток крови (ККК) и их особенности при разных типах анемий
Происхождение нормальных красных клеток крови (ККК) и их особенности при разных типах анемий
При соответствующей стимуляции большие количества этих клеток формируются из стволовых клеток КОЕ-Э.
Сразу после образования проэритробласт делится много раз, в результате формируются много зрелых красных клеток крови. Клетки первой генерации называют базофилъными эритробластами, поскольку они окрашиваются основными красителями; к этому времени клетка накапливает очень мало гемоглобина.
В последующих генерациях, как показано на рисунке, клетки наполняются гемоглобином до концентрации, равной примерно 34%, ядро сжимается до небольшого размера, и его конечный остаток абсорбируется или выдавливается из клетки. В то же время абсорбируется эндоплазматический ретикулум. На этом этапе клетку называют ретикулоцитому поскольку она все еще содержит небольшое количество базофильного материала, состоящего из остатков аппарата Гольджи, митохондрий и некоторых других цитоплазматических органелл.
Во время этой ретикулоцитарной стадии клетки выходят из костного мозга в кровеносные капилляры путем диапедеза (просачиваясь сквозь поры капиллярной мембраны).
Остаточный базофильный материал ретикулоцита в норме исчезает в течение 1-2 сут, и после этого клетка становится зрелым эритроцитом. В связи с короткой жизнью ретикулоцитов их концентрация среди всех клеток крови обычно немного ниже 1%.
Регуляция продукции красных клеток крови. Роль эритропоэтина
Общая масса эритроцитов в системе кровообращения регулируется в узких пределах, поэтому, с одной стороны, (1) всегда имеется адекватное количество красных клеток крови для обеспечения достаточного транспорта кислорода из легких к тканям, с другой стороны, (2) клеток не становится так много, чтобы они мешали кровотоку. Известные в настоящее время данные об этом контроле представлены в виде схемы на рисунке ниже и заключаются в следующем.
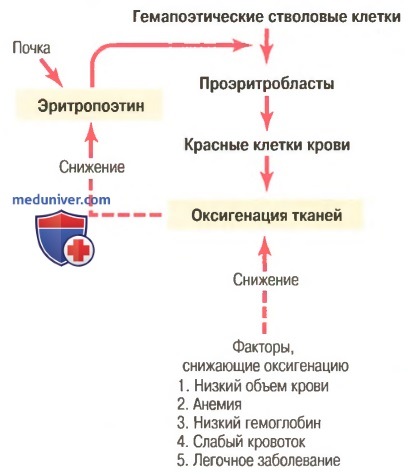 Увеличение продукции красных клеток крови, связанное с функцией механизма эритропоэтина при снижении оксигенации тканей
Увеличение продукции красных клеток крови, связанное с функцией механизма эритропоэтина при снижении оксигенации тканей
а) Оксигенация тканей является наиболее важным регулятором продукции красных клеток крови. Любое состояние, ведущее к уменьшению доставки кислорода тканям, обычно увеличивает скорость образования эритроцитов. Следовательно, если в результате кровотечения или любого другого состояния у человека развивается анемия, костный мозг немедленно усиливает продукцию красных клеток крови. Кроме того, разрушение значительной части костного мозга по любой причине, особенно при рентгенотерапии, ведет к гиперплазии оставшегося костного мозга, что отражает попытку удовлетворить потребности организма в эритроцитах.
На очень больших высотах, где количество кислорода в воздухе сильно снижено, снабжение тканей кислородом становится недостаточным, и продукция красных клеток крови резко возрастает. В этом случае образование эритроцитов регулирует не их концентрация в крови, а несоответствие между доставкой кислорода тканям и потребностями тканей в кислороде.
Скорость продукции красных клеток крови возрастает также при различных болезнях системы кровообращения, сопровождаемых снижением кровотока через периферические сосуды, особенно на фоне недостаточности поглощения кислорода при прохождении крови через легкие. Так, тканевая гипоксия, характерная для хронической сердечной недостаточности и многих легочных заболеваний, стимулирует продукцию красных клеток крови, что ведет к увеличению гематокрита и, как правило, общего объема крови.
б) Эритропоэтин стимулирует продукцию красных клеток крови, а его образование возрастает в ответ на гипоксию. Главным стимулом для продукции эритроцитов при состояниях с низким содержанием кислорода является циркулирующий в крови гормон эритропоэтин — гликопротеин с молекулярной массой около 34000. При отсутствии эритропоэтина гипоксия практически не стимулирует продукцию эритроцитов. Но когда система эритропоэтина функционирует, гипоксия вызывает заметное увеличение секреции эритропоэтина, а он, в свою очередь, усиливает образование красных клеток крови до тех пор, пока гипоксия не исчезнет.
в) Роль почек в формировании эритропоэтина. У здорового человека примерно 90% всего эритропоэтина синтезируется в почках; остальная часть образуется в основном в печени. Пока точно не известно, где именно в почках синтезируется эритропоэтин. Одним из возможных мест секреции эритропоэтина являются эпителиальные клетки почечных канальцев, поскольку анемичная кровь околоканальцевых капилляров не способна удовлетворить их высокую потребность в кислороде, что стимулирует продукцию эритропоэтина.
Иногда секрецию эритропоэтина почками стимулирует гипоксия не в почках, а в других частях тела, что позволяет предположить возможное существование некоего внепочечного сенсора, который посылает к почкам дополнительный сигнал, стимулирующий продукцию этого гормона. В частности, норадреналин, адреналин и некоторые из простагландинов стимулируют продукцию эритропоэтина.
Когда у человека удаляют обе почки или если они разрушены болезнью, у человека неминуемо развивается тяжелая анемия, поскольку эритропоэтин, синтезируемый другими тканями (в основном печенью), составляет лишь 10% нормы и может обеспечить формирование не более половины красных клеток крови, необходимых организму.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»



