Как называется скопление нейронов разных размеров и различной формы разделенных множеством волокон
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
Микроскопически кора имеет полосчатый, или пластинчатый, вид, отражающий распределение ее клеток и нервных волокон, а также веерообразную организацию клеточных элементов. Общая цитоархитектоника (схема строения, основанная на типе клеток; схему строения в зависимости от распределения миелинизированных волокон обозначают термином миелоархитектоника) во многом варьирует от зоны к зоне, что позволяет составить «карту» коры с десятками гистологически различных зон. Больших успехов удалось достичь в соотнесении этих зон со специфическими функциями. Несмотря на то, что это имеет теоретическую ценность, подобное разделение довольно упрощено, так как часто принимают во внимание только ключевые функции более обширных функциональных систем, взаимодействующих с другими отделами мозга.
а) Пластинчатое строение. Пластинчатое распределение нейронов характерно для всех отделов коры. Филогенетически «старые отделы», включая древнюю кору (обонятельную кору) и старую кору (гиппокампальная формация и зубчатое ядро; участвуют в процессах памяти) образованы тремя клеточными пластинками. Однако в новой коре (неокортексе или изокортексе, название которой указывает на единый нейрогенез коры, закончившийся образованием шестислойного строения), составляющей оставшиеся 90% (подавляющую часть коры больших полушарий), происходит переход к шестислойному строению.
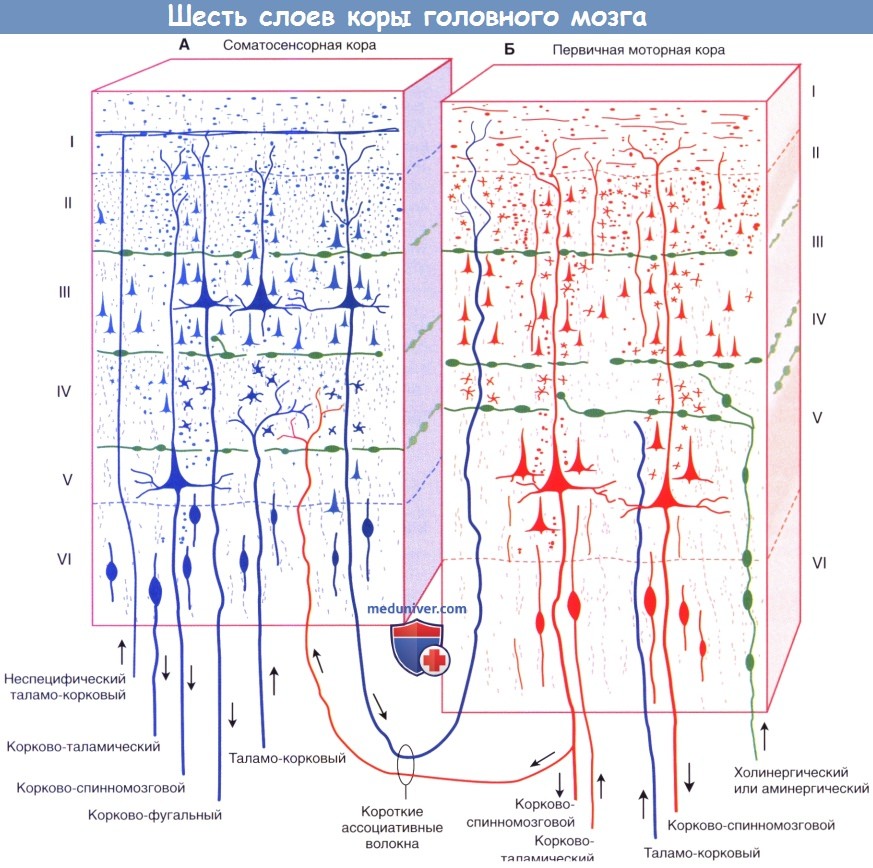 Шестислойное строение изокортекса коры головного мозга.
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
б) Колончатое строение. Помимо хорошо различимой пластинчатой организации коры больших полушарий, можно наблюдать также лучистую, или «колончатую», организацию клеточных структур. Колончатое строение новой коры стало основой исследования функционирования групп нейронов в соматосенсорной коре животных. Подобные лучевидно расположенные группы клеток, вероятно, представляют собой отдельные зоны с одинаковыми физиологическими свойствами и образуют структурную основу для создания более сложных функций. Группы колонн могут образовывать модули, участвующие в обработке различных сенсорных модальностей и функций.
Установлено, что колонны в различных отделах коры имеют разное строение, обусловленное вариабельностью многочисленных свойств, включая строение и число клеток, онтогенез, синаптические связи и молекулярные маркеры. Все это влияет на различные функциональные характеристики и реакции в ответ на возбуждение. В качестве организующего принципа подобная концепция колончатого строения представляет определенную пользу, однако удобнее рассматривать кору как совокупность горизонтального (пластинчатого) и вертикального (лучистого) типов строения. Несмотря на то, что колонны не представляют собой отдельные подобные образования с четкими границами, такая концепция более точно отражает строение коры, а также наблюдаемые экспериментально функциональность, «экономию пространства» и пластичность коры.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
Лежащая в основе организации коры «схема» приводит к тому, что каждая из колонн становится модально (функционально) специфичной после «обработки» информации отдельными элементами. Однако конечная реакция проекционных нейронов колонн может значительно варьировать в зависимости от степени их возбуждения и информации, подходящей к каждому нейрону. Например, одна колонна может реагировать на движение в определенном суставе, но не на раздражение кожи над ним; однако при изменении условий их реакция также может изменяться.
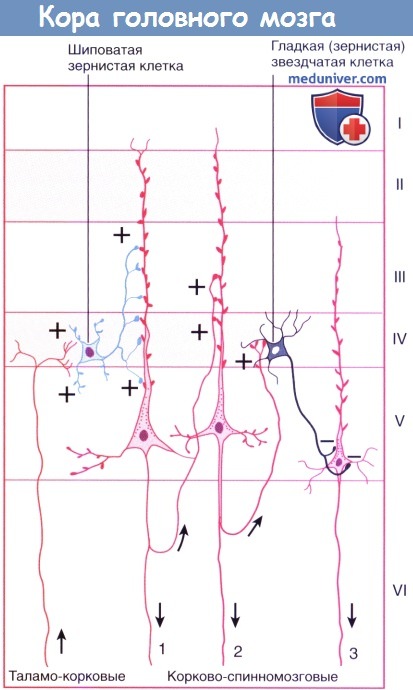 Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Пирамидные клетки имеют пирамидоподобную форму с вершиной, направленной к поверхности. Размеры клеток в высоту составляют 20-30 нм в слоях II и III и более чем в два раза больше в слое V. Самые крупные — гигантские пирамидные клетки Беца — расположены в моторной коре. Единственный апикальный дендрит каждой пирамидной клетки достигает слоя I, заканчиваясь на пучке дендритов. Несколько базальных дендритных ветвей отходят от базальных «углов» клетки и веерообразно расходятся к соответствующим слоям. Апикальные и базальные дендритные ветви свободно разветвляются и усыпаны дендритными шипиками. Большинство пирамидных клеток расположено в слоях II-III и V-VI. Отходящие от основания клеток аксоны отдают несколько возвратных ветвей до вхождения в подлежащее белое вещество, их функция — возбуждение соседних пирамидных клеток.
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Нейромедиатором различных по структуре и классификации непирамидных тормозных вставочных нейронов служит ГАМК. [Сложная классификация нейронов новой коры постоянно меняется. Гладкие звездчатые (зернистые) клетки можно обнаружить во всех слоях коры; их дендриты веерообразно расходятся во всех направлениях, а их аксоны образуют локальные древовидные разветвления, поэтому их иногда называют нейронами локальных сплетений. Несмотря на их уникальные морфологические характеристики, нейроглиеформные, клетки-канделябры и корзинчатые клетки считают специализированными типами зернистых клеток. Наш совет: если Вы встречаете термины «зернистая» или «гладкая звездчатая» клетка, для облегчения чтения и понимания их следует рассматривать как вставочные нейроны.]
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
Корзинчатые клетки лежат преимущественно в слоях II и V, а свое название получили благодаря тому, что их аксоны формируют околоклеточные «корзинки» вокруг тел пирамидных клеток, дистальных отделов их дендритов и аксонов других корзинчатых клеток.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
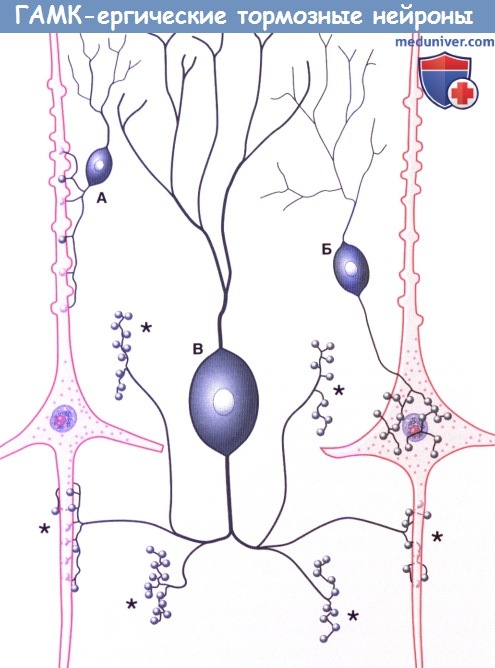 Три морфологических типа ГАМК-ергических тормозных нейронов:
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Редактор: Искандер Милевски. Дата публикации: 22.11.2018
Ретикулярная формация. В срединной части ствола мозга расположена ретикулярная формация (РФ) — скопление нейронов разных размеров и формы
В срединной части ствола мозга расположена ретикулярная формация (РФ) — скопление нейронов разных размеров и формы, разделенных множеством проходящих в разных на правлениях волокон, напоминающих сеть (лат. reticulum). В РФ локализовано большое количество нейронов различного вида и размера, сгруппированных в ядра.
Общие черты нейронов РФ — это форма и характер организации их связей. Нейроны РФ являются клетками типа Гольджи I (с длинными аксонами). При этом аксоны имеют две ветви, идущие рострально и каудально. Таким образом, от клеток РФ начинаются как восходящие, так и нисходящие пути, дающие многочисленные коллатерали, окончания которых образуют синапсы на нейронах всех мозговых уровней, т.е. один ретикулярный нейрон может посылать генерируемые им импульсы одновременно в различные структуры ЦНС (рис. 27)
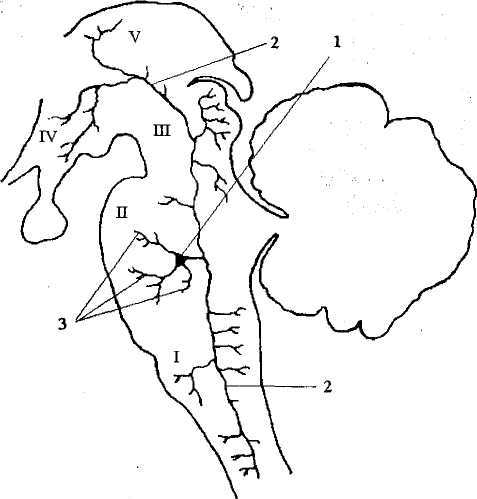
Рис. 27. Нейрон ретикулярной формации и разветвления его аксона:
1 — тело нейрона; 2 — аксон; 3 — дендриты; I — продолговатый мозг;
II— Варолиев мост; III — средний мозг; IV— гипоталамус;
Длинные ветвящиеся дендриты нейронов РФ ориентированы преимущественно в плоскости, перпендикулярной про дольной оси мозга. Для РФ характерна конвергенция (схождение) афферентации от разных сенсорных систем на одном нейроне. Например, на одной ретикулярной клетке могут образовывать синапсы чувствительные волокна, несущие ин формацию от кожных, зрительных и слуховых рецепторов.
В связи с такими особенностями связей (как афферентных, так и эфферентных) ретикулярная система была названа не специфической в отличие от специфических систем, которые принимают информацию от совершенно определенных структур и посылают ее по конкретным «адресам».
По структурно-функциональным критериям РФ делится на 3 зоны: медианную — по средней линии, медиальную — внутренние отделы ствола и латеральную, нейроны которой лежат вблизи сенсорных ядер. В медиальных отделах РФ продолговатого мозга и моста находят большие и даже гигантские нейроны, в латеральных отделах этого же уровня обнаруживают малые и средние нейроны; в среднем мозгу расположены преимущественно малые нейроны. Медианная зона протянута от продолговатого мозга до каудальных (задних) отделов среднего мозга. Структуры, входящие в эту зону, объединяются под общим названием ядра шва. В среднем мозгу к ядрам шва примыкают ядра центрального серого вещества, по ряду особенностей сходные с ядрами РФ. Для нейронов ядер шва характерно присутствие в качестве медиатора серотонина.
Основной объем афферентации РФ получает от сенсорных образований, таких как чувствительные ядра, спинно-ретикулярный тракт и т.п. Вместе с тем на нейронах РФ так же образуют синапсы коллатерали от ряда нисходящих путей, в частности кортикоспинального и руброспинального трактов. Получает РФ афференты и от мозжечка (от ядер шатра).
Эфференты РФ образуют две основные системы волокон — восходящую и нисходящую. Восходящие аксоны идут в передний мозг— к неспецифическим ядрам таламуса (про межуточный мозг), коре больших полушарий; нисходящие аксоны направляются в СМ. Кроме того, волокна от РФ идут к мозжечку.
Многочисленные связи существуют и внутри РФ между различными ее образованиями, а также между ядрами РФ и другими стволовыми структурами.
РФ — мозговая система, регулирующая работу ЦНС и выполняющая важнейшие интегративные (объединяющие) функции. Эти функции очень многочисленны, хотя и не до конца исследованы. РФ играет ключевую роль в управлении общим уровнем активности нервной системы, в частности в регуляции цикла «сон— бодрствование». Через пути, связывающие РФ со спинным мозгом, она принимает участие в управлении позой, локомоцией и целенаправленными движениями. Ядра РФ участвуют также в регуляции, связанной с жизненно важными рефлексами. Так, в РФ продолговатого мозга и моста находятся центры дыхания (с подразделением на центр вдоха и центр выдоха), сосудодвигательный центр (регулирующий тонус сосудов и работу сердца), центр слюноотделения и выделения других пищеварительных соков, центр глотания, а также центры таких защитных рефлексов, как кашель, чихание, рвота.
Из-за наличия в РФ дыхательного и сосудодвигательного центров нормальная работа этого отдела жизненно необходима. В то время как повреждение, например, структур конечно го мозга нередко почти не вызывает последствий в связи с большими компенсаторными возможностями ЦНС, даже не значительные повреждения РФ мозгового ствола приводят к тяжелым нарушениям функций организма, и даже к смерти.
Мозжечок
Общее строение
Мозжечок, cerebellum, управляет точными согласованными движениями и сохранением равновесия. Его ширина равняется примерно 10 см, толщина — 3 см. Масса мозжечка составляет около 11% от массы всего головного мозга. Сверху он покрыт корой, под которой находится белое вещество. В толще белого вещества лежат ядра серого вещества. Такую же организацию имеют и большие полушария. Поэтому мозжечок называют иногда «малый мозг».
Мозжечок расположен на задней стороне ствола и состоит из двух полушарий, hemispheria, и непарной соединительной части — червя, vermis. Нижняя поверхность червя является крышей IV желудочка. Сверху мозжечок покрыт большими полушариями переднего мозга. Мозжечок имеет три пары ножек— нижние, средние и верхние. Они представляют собой волокна белого вещества, соединяющие мозжечок с другими мозговыми структурами. На разрезе белое вещество мозжечка имеет вид мелких веточек. В результате общая картина белого и серого вещества напоминает дерево (ее даже называют «древом жизни»).
Кора мозжечка образует узкие извилины — листки мозжечка, отделенные друг от друга бороздами. Крупные борозды делят мозжечок на доли и дольки (рис. 28). Каждой дольке червя соответствует долька полушария (см. таблицу).
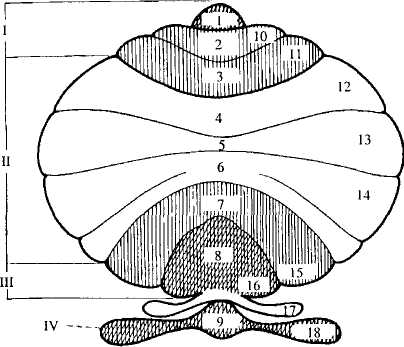
Рис. 28. Схема мозжечка, показывающая его доли и дольки:
1— передняя доля, II— средняя доля, III — задняя доля, IV— флоккуло-нодулярная доля; 1-9 — дольки червя: 1 — язычок мозжечка, 2 — центральная долька, 3 — верхушка, 4 — скат, 5— листок, 6— бугор, 7— пирамида, 8— язычок червя, 9 — узелок, 10—18— дольки полушарий: 10 — крыло центральной дольки,
11 — передняя и 12 — задняя части четырехугольной дольки,
13— верхняя и 14— нижняя полулунные дольки, 15 — двубрюшная
долька, 16— миндалина, 17— околоклочок, 18 — клочок
| Доля | Дольки | мозжечка | ||
| мозжечка | Червь | Полушария | ||
| Верхняя доля | 1) | язычок мозжечка | 1) | связка язычка |
| 2) | центральная долька | 2) | крылья центральной дольки | |
| 3) | горка: | i) | четырехугольная долька: | |
| а) | верхушка | а) | передняя часть | |
| б) | скат | б) задняя часть | ||
| Задняя доля | 4) | листок | 4) | верхняя полулунная долька |
| 5) | бугор | 5) | нижняя полулунная долька | |
| Нижняя доля | 6) | пирамида | 6) | двубрюшная долька |
| 7) | язычок червя | 7) | миндалина | |
| 8) | узелок | 8) | клочок |
Ядра мозжечка хорошо видны на его горизонтальном разрезе (рис. 29). Наиболее медиально в черве лежат ядра шатра, несколько латеральнее — шаровидные ядра, затем — пробковидные ядра. Наиболее латерально в центре полушарий находятся зубчатые ядра, каждое из которых своей формой напоминает складчатую чашу.
У различных частей мозжечка разный филогенетический возраст, в связи с чем выделяют древний, старый и новый мозжечок.
К древнему мозжечку, archicerebellum[1], относится небольшая долька, расположенная в наиболее каудальной части полушарий — клочок, flocculus, и соответствующий ей в черве узелок, noclulus. Сюда же относят ядра шатра. Древний мозжечок называют иногда клочково-узелковой или флоккуло-нодулярной долей (см. рис. 28). Он связан с вестибулярными ядрами, поэтому при его повреждении нарушается способность поддерживать равновесие тела, как и при патологиях вестибулярного аппарата. Походка таких больных становится шатающейся, они часто страдают головокружением, тошнотой и рвотой.
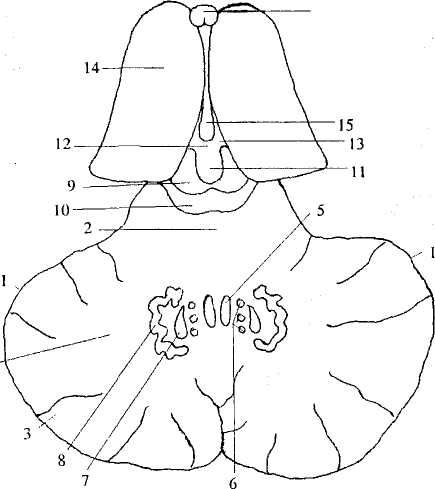
1 — полушарие мозжечка; 2 — червь; 3 — кора мозжечка;
4 — белое вещество мозжечка; 5 — ядро шатра; 6 — шаровидное
ядро; 7 — пробковидное ядро; 8 — зубчатое ядро; 9 — верхний холмик
четверохолмия; 10— нижний холмик четверохолмия; 11 — эпифиз; 12 —
комиссура поводков; 13 — треугольник поводков; 14 — таламус; 15 — III
мозговой желудочек; 16— колонны свода
К старому мозжечку, paleocerebellum, относят весь червь, кроме узелка и ската, переднюю долю полушарий, а также шаровидное и пробковидное ядра. В связи с выполнением общей функции их объединяют в промежуточное ядро мозжечка. Старый мозжечок образовался в связи с развитием способности к локомоции. При его поражении нарушается работа мускулатуры шеи и туловища, затруднено поддержание позы. Кроме того, нарушается точность и сила сгибательно-разгибательных движений конечностей (как правило, наблюдается их патологическое усиление). Наиболее известный тест на наличие повреждений старого мозжечка — пальце-носовая проба. Основные входные влияния поступают в старый мозжечок по спинно-мозжечковым трактам; основные выходы идут в РФ и крупноклеточную часть красно го ядра.
Большая часть полушарий и зубчатое ядро образуют самую молодую часть — новый мозжечок, neocerebellum. Он развивается в связи с совершенствованием тонкой моторики конечностей. Особенно сильное развитие новый мозжечок получил у человека в связи с чрезвычайно дифференцированными движениями при трудовых процессах. Поражение этой части отражается более всего на движениях кистей конечностей, в которых развивается атаксия — нарушение координации и точности движений, так, например, резко ухудшается почерк. Может также изменяться и речь — она становится растянутой, замедленной или взрывчатой.
Новый мозжечок через зубчатые ядра и таламус в промежуточном мозгу связан с двигательной корой больших полушарий. В результате он способен регулировать активность кортикоспинального тракта и управлять такими сложнейшими двигательными навыками как, например, письмо, печатание на клавиатуре, игра на музыкальных инструментах и т.п. Он участвует в двигательном обучении и управлении наиболее сложными движениями, в частности движениями пальцев.
Таким образом, основной функцией мозжечка является регуляция и коррекция движений в процессе их выполнения, программирование движений и двигательное научение, т.е. перевод произвольных движений в автоматизированные. За поминание параметров двигательных реакций в мозжечке происходит за счет того, что «копии» всех программ произвольных движений (т.е. движений, контролируемых сознанием) через мелкоклеточную часть красного ядра и ядра оливы поступают на клетки Пуркинье (см. далее). При поражении полушарий мозжечка нарушается точность, согласованность и быстрота выполнения движений, не вырабатываются новые тонкие двигательные навыки, нарушаются речь и письмо, наблюдается дрожь в конечностях.
Кора мозжечка
В коре мозжечка различают пять основных типов нейронов, причем все нейроны за исключением клеток-зерен — тормозные, т.е. их аксоны образуют на других клетках синапсы, под действием которых постсинаптические нейроны ослабляют свою активность.
Рассмотрим строение коры мозжечка более подробно (рис. 30). Начнем со среднего (ганглиозного) слоя. Он состоит из клеток Пуркинье. Это крупные (диаметром
Аксоны клеток Пуркинье идут вглубь мозжечка в белое вещество и заканчиваются на клетках ядер мозжечка. Необходимо обратить внимание на то, что только аксоны клеток Пуркинье являются эфферентами коры мозжечка.
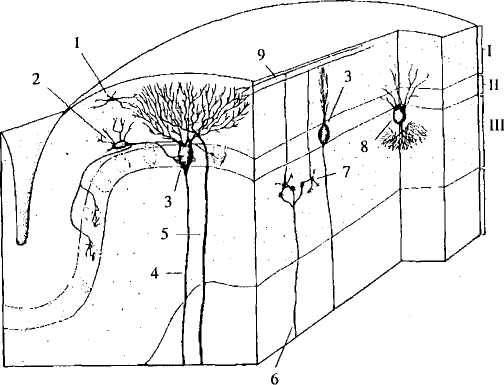
Рис. 30. Кора мозжечка:
I— молекулярный слой, II— ганглиозный слой, III — зернистый слой,
IV— белое вещество, 1 — звездчатая клетка; 2 — корзинчатая клетка,
3 — клетка Пуркинье; 4 — аксон клетки Пуркинье; 5 — лазающее
волокно; 6 — мшистое волокно; 7 — зернистая клетка; 8 — клетка
Гольджи; 9— параллельное волокно
Кроме зернистых клеток, во внутреннем слое коры мозжечка есть клетки Гольджи. Их дендриты поднимаются в молекулярный слой, где контактируют с параллельными волок нами. Аксоны этих клеток образуют тормозные синапсы на клетках-зернах.
В наружном (молекулярном) слое находятся дендриты клеток Пуркинье и клеток Гольджи, а также аксоны клеток-зерен (параллельные волокна). Кроме того, здесь есть два типа тормозных нейронов — корзинчатые и звездчатые.
Корзинчатые нейроны лежат в нижней части молекулярно го слоя. Это мелкие (10-15 мкм) клетки с тонкими длинными дендритами. Их аксоны идут параллельно поверхности коры. От аксонов отходят коллатерали, которые оплетают тела клеток Пуркинье, образуя характерную систему корзинок.
Звездчатые клетки лежат выше корзинчатых. Их аксоны образуют одиночные синапсы на телах и дендритах клеток Пуркинье.
Афференты коры мозжечка образуют две системы волокон — лазающие (лиановидные) и мшистые (моховидные).
Лазающие волокна — это аксоны нейронов, лежащих в ядрах олив (см. 7.2.2). Они оканчиваются на соме и дендритах клеток Пуркинье. На каждой клетке Пуркинье образует синапсы только одно лазающее волокно.
Мшистые волокна, которых гораздо больше, чем лазающих, образуют синапсы на дендритах клеток-зерен и приходят от самых разных структур ЦНС (см. далее). Одно мшистое волокно образует синапсы примерно на 20 клетках-зернах.
Система афферентных волокон организована так, что по ступающая в кору мозжечка импульсация в конечном итоге адресуется клеткам Пуркинье. При запуске движения происходит торможение клеток Пуркинье через, прежде всего, звездчатые и корзинчатые клетки. Как следствие, на время прекращается тормозное действие аксонов клеток Пуркинье на ядра мозжечка. В результате наблюдается активация тех двигательных программ, рефлекторные дуги которых проходят через соответствующие нейроны ядер.
Основные связи внутри мозжечка, а также между мозжечком и другими структурами ЦНС представлены на рис. 31.



