Механизмы миофибриллярной гипертрофии скелетных мышц человека
В статье рассмотрены механизмы миофибриллярной гипертрофии скелетных мышц человека с позиций ряда медико-биологических дисциплин: анатомии, гистологии, цитологии, биохимии, физиологии и спортивной медицины. В публикации анализируются различные гипотезы, объясняющие механизмы повышенного синтеза белка в мышцах под воздействием тренировки силовой направленности. В настоящее время наиболее распространена гипотеза механического повреждения мышечных волокон.
Самсонова А.В. Механизмы миофибриллярной гипертрофии скелетных мышц человека //Мат. 1 Всерос. Конф. С межд. Участием «Совершенствование учебного процесса в условиях современного вуза».- Белгород: БегГУ, 2012.- С. 388-395
Самсонова А.В.
МЕХАНИЗМЫ ГИПЕРТРОФИИ СКЕЛЕТНЫХ МЫШЦ ЧЕЛОВЕКА
ВВЕДЕНИЕ
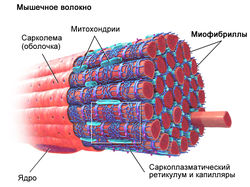
Миофибриллярная гипертрофия – адаптация скелетных мышц человека к силовым нагрузкам при направленности тренировочного процесса на увеличение их объема или силы. Установлено, что при этом типе гипертрофии возрастает количество и объем миофибрилл – основных элементов мышечного волокна [2, 10, 22].
Целью исследования являлась разработка концепции, описывающей механизмы миофибриллярной гипертрофии скелетных мышц человека под воздействием нагрузок силовой направленности.
РЕЗУЛЬТАТЫ
В настоящее время существует несколько гипотез, объясняющих повышенный синтез белка в скелетных мышцах человека.
В основе первой гипотезы – энергетической [2, 10, 22] – лежит предположение о том, что нарушение равновесия между потреблением и восстановлением основной энергетической «валюты» – АТФ – стимулирует процессы, протекающие в мышцах, в результате чего происходит их гипертрофия. Известно, что содержание в мышце АТФ ограничено. При проведении интенсивных силовых тренировок в мышцах возникает недостаток АТФ, что является для организма серьезным предупреждающим сигналом. Недостаток АТФ неблагоприятно сказывается на метаболизме белка. Следует отметить, что в результате интенсивных силовых тренировок происходит также большое разрушение белков мышц. Расходуются не только миофибриллярные белки, но и ферменты и гормоны (функциональные белки), которые играют важную роль в сокращении мышц. Известно, что белки состоят из аминокислот. Основой аминокислот является азот. Установлено, что силовые тренировки приводят к выделению большого количества азота в виде продуктов распада мышечных белков (мочевина). Во время напряженных силовых тренировок и непосредственно после них распад белка значительно превосходит его восстановление. По мнению [10], это связано, в первую очередь, с нехваткой АТФ. Таким образом, равновесие между постоянно протекающими процессами распада и восстановления, которое наблюдается в нормальных условиях, серьезно нарушается. В последующих фазах восстановление белковых структур с помощью пищи, богатой протеинами, осуществляется настолько интенсивно, что их количество превышает исходный уровень за счет явления суперкомпенсации. Вследствие этого увеличивается площадь поперечного сечения миофибрилл. Повторные интенсивные тренировки воздействуют уже на большую площадь миофибрилл. Таким образом, интенсивные силовые тренировки становятся менее опасными для организма. Помимо увеличения площади поперечного сечения миофибрилл и их количества, в мышечных волокнах происходит заметное увеличение запасов фосфатных соединений, богатых энергией. Таким образом, организм приспосабливается к нагрузке.
Более подробно о механизмах гипертрофии мышц Вы можете прочесть в моей книге «Гипертрофия скелетных мышц человека«
В основе второй гипотезы [8, 9] лежит предположение о том, что пусковым стимулом синтеза белка в мышцах является их ацидоз[1], вызванный накоплением в мышцах кислых продуктов метаболизма (ионов водорода), а также увеличение содержания в мышечных волокнах креатина.
Схема повышенного синтеза белка выглядит следующим образом. В ходе выполнения силовых упражнений с большими отягощениями (до 80% от максимума) энергия АТФ тратится на выполнение механической работы. Ресинтез АТФ начинается благодаря запасам креатинфосфата в мышечном волокне. В результате чего в мышечном волокне появляется креатин. Накопление креатина в саркоплазматическом пространстве служит мощным эндогенным стимулом, возбуждающим белковый синтез в скелетных мышцах. Показано, что между содержанием сократительных белков и содержанием креатина имеется строгое соответствие. Креатин, видимо, воздействует на синтез иРНК (информационной РНК), т.е. на процессы транскрипции в ядрах мышечных волокон.
Креатин активизирует деятельность всех метаболических путей, связанных с образованием АТФ (гликолиз в саркоплазме, аэробное окисление в митохондриях). Так как мощность гликолиза меньше скорости потребления АТФ, а мощность аэробного окисления еще ниже, чем гликолиза, в клетке начинают накапливаться ионы водорода, лактат и АДФ. Повышение концентрации ионов водорода вызывает лабилизацию мембран (увеличение размеров пор в мембранах, что ведет к облегчению проникновения гормонов в клетку), активизирует действие ферментов, облегчает доступ гормонов к наследственной информации, к молекулам ДНК. В ответ на одновременное повышение концентрации креатина и ионов водорода в ядрах клетки интенсивнее образуется РНК.
В основе третьей гипотезы лежит предположение, что пусковым стимулом для возрастания синтеза белка в мышцах является гипоксия[2]. Такое предположение связано с тем, что при выполнении упражнений силовой направленности при напряжении мышцы более 60% от максимума, капилляры и артериолы мышцы сдавливаются, и кровь к сокращающимся мышцам не поступает [22].
Гипоксия, развивающаяся в мышцах в процессе нагрузки, ведет к накоплению кислых метаболитов и закислению саркоплазмы. Калиемия, вызывающая сужение кровеносных сосудов, усиливает состояние гипоксии. Энергетические ресурсы ткани истощаются. Изменение энергетического метаболизма проявляется в нарушении транспорта ионов через мембраны клеток, повышении концентрации кальция и активации протеолиза[3]. В первую очередь, активируются калпаины – нелизосомальные протеазы, которые играют важную роль в запуске расщепления белков скелетных мышц, воспалительных изменениях и процессе регенерации. После окончания выполнения физических упражнений за гипоксией следует реперфузия[4]. Доказано, что интенсивные физические нагрузки вызывают сильную метаболическую гипоксию мышц, последствия которой после прекращения нагрузки оказываются сходными с последствиями реперфузии при ишемии. Приток кислорода в мышцы остается на высоком уровне, хотя метаболический запрос ткани в отношении кислорода снижается. Это вызывает активацию процессов перекисного окисления липидов (ПОЛ), что нарушает целостность сарколеммы мышечного волокна, повреждение и деградацию его сократительных белков, а также белков цитоскелета [11]. Одновременно с этим в мышечном волокне развиваются воспалительные процессы, что выражается в повышении содержания лейкоцитов в скелетных мышцах через 24 часа после тренировки. Каскад процессов, происходящих в мышце после тренировки, приводит к появлению запаздывающих болевых ощущений и ухудшению функционального состояния мышцы, что выражается в уменьшении уровня максимальной силы. Затем в поврежденных мышечных волокнах активируются клетки-сателлиты, которые активно участвуют в регенерации мышцы и восстановлении ее функциональной активности [4, 5].
В основе четвертой гипотезы, которая получила в настоящее время широкое распространение, лежит предположение о том, что пусковым стимулом для возрастания синтеза белка в мышцах является механическое повреждение мышечных волокон и миофибрилл, после которого следует их регенерация.
Доказано, что после больших физических нагрузок происходит повреждение мышечных волокон [12, 15, 3]. Д.Дж. Нейман с соавт. [20] показали, что сразу после физических упражнений 16% мышечных волокон имели легкие повреждения, 16% – более сильные и 8% — очень сильные. Кроме того, эти авторы утверждали, что повреждения, замеченные немедленно после выполнения упражнения, были предшественниками более сильных повреждений, которые отмечались в последующих биопсиях. Ж.Фрайден с соавт. [13] показали, что через час после выполнения эксцентрических упражнений у человека в 32% мышечных волокон были обнаружены повреждения, а через три дня повреждения были обнаружены в 52% мышечных волокон. М.Джибала с соавт. [15] установили, что даже однократная высокоинтенсивная силовая тренировка приводит к повреждению большого количества мышечных волокон (от 30 до 80%). При этом более сильные повреждения обнаруживаются в волокнах II типа по сравнению с волокнами I типа [13]. Установлено также, что волокна II типа повреждаются в первую очередь [16].
Из компонентов мышечного волокна сильные повреждения наблюдаются в сарколемме, саркоплазматическом ретикулуме, миофибриллах, цитоскелете. Наиболее подверженными разрушению оказываются Z-диски мышечного волокна [12, 14, 15]. Если повреждается сарколемма мышечного волокна, в крови появляются ферменты, содержащиеся в саркоплазме. Очень часто в крови обнаруживается фермент креатинкиназа, который участвует в креатинфосфатном пути ресинтеза АТФ. Доказано, что содержание в крови ферментов после значительных нагрузок силовой направленности может увеличиваться в 100 раз [3]. Показано, что повреждение мышечных волокон различных типов можно диагностировать посредством определения в сыворотке крови легкой и тяжелой изоформ миозина. При изучении различной степени повреждения мышцы установлено, что при самых легких повреждениях мышечных волокон уровень легкой изоформы миозина увеличивается с 625 мг/л до 2880 мг/л, то есть более чем в 4 раза [16]. Появление в крови легкой изоформы миозина свидетельствует о повреждении мышечных волокон II типа. Повреждение мышечных волокон сопровождается запаздывающими болевыми ощущениями (DOMS). Повреждение мышечных волокон и миофибрилл запускает процессы регенерации мышечной ткани (рис.1).

Рис. 1. Схематическое повреждение мышечного волокна [19]. Обозначения: А – напряжение мышцы ведет к повышению концентрации ионов кальция (черные кружки); В – повышение концентрации ионов кальция приводит к активации калпаинов и избирательному повреждению элементов цитоскелета; С – последующая активность мышцы повреждает цитоскелет.
Если мышечное волокно было разорвано или повреждено, в поврежденном участке образуется некротическая зона. При этом на некотором расстоянии от места травмы возникает полное разрушение сарколеммы, саркоплазмы и органелл, хотя за пределами этой зоны волокно сохраняет свою жизнеспособность. Считается, что этот процесс инициируется увеличенным количеством внутриклеточного кальция (Са 2+ ), который поступает в саркоплазму мышечного волокна из поврежденного саркоплазматического ретикулума. Ионы кальция активируют ферменты – протеазы, которые расщепляют белки в миофибриллах. В первую очередь, активируются калпаины – протеолитические ферменты, которые воздействуют на белки цитоскелета [14, 16]. Именно белки цитоскелета разрушаются в первую очередь [19, 14]. Появление в волокне обрывков белковых молекул активирует лизосомы, переваривающие с помощью содержащихся в них ферментов белковые структуры, которые необходимо уничтожить. Если лизосомы не справляются с объемом работы, то через сутки активируются более мощные чистильщики – фагоциты. Фагоциты – клетки, находящиеся в крови и тканевой жидкости. Основная их задача – уничтожение поврежденных тканей и чужеродных микроорганизмов. Фагоциты проникают в волокно, потребляют его содержимое и выводят остатки. Именно продукты жизнедеятельности фагоцитов вызывают воспалительные процессы в мышцах через сутки после тренировки. В это же время в мышечном волокне начинается процесс его «ремонта». С гистологической точки зрения, при регенерации возможно не только восстановление целостности поврежденных мышечных волокон, но и возникновение новых мышечных волокон [1, 3]. Регенерация идет тем интенсивнее, чем больше освобождается из-под базальной мембраны клеток-сателлитов.
Иногда сильные повреждения обнаруживаются в миофибриллах, хотя внешне мышечное волокно сохраняет свою целостность. Некоторые саркомеры могут быть более растянутыми по сравнению с другими. Это означает, что поврежден цитоскелет мышечного волокна, то есть продольные филаменты, связывающие соседние саркомеры в одной миофибрилле, а также поперечные филаменты, связывающие соседние миофибриллы между собой и с сарколеммой. Повреждение цитоскелета неизбежно приводит к незначительным повреждениям сарколеммы и как следствие – активации деятельности клеток-сателлитов, которые в мышечном волокне располагаются между сарколеммой и базальной мембранной.
Многочисленными исследованиями доказано, что в результате выполнения силовых упражнений, в первую очередь, повреждаются Z-диски, соединяющие саркомеры друг с другом в миофибрилле. Напомним, что к Z-дискам прикрепляются тонкие филаменты. Считается, что Z-диски являются «слабым звеном» миофибриллы. При этом возможен как полный разрыв миофибрилл в области Z-дисков так и растягивание Z-дисков с сохранением целостности миофибриллы (рис. 2).

ВЫВОДЫ
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
[1] Ацидоз – закисление внуренней среды организма, связано с накоплением в тканях кислых продуктов обмена веществ. При напряженной мышечной работе ацидоз приводит к развитию утомления.
[3] Протеолиз – распад белков, приводящий к образованию аминокислот.
[4] Реперфузия – возобновление притока кислорода к тканям.
Гипертрофия мышц
Содержание
Механизмы гипертрофии скелетных мышц [ править | править код ]

Гипертрофия скелетных мышц (греч. hyper – больше и греч. trophe – питание, пища) – это адаптационное увеличение объема или массы скелетной мышцы. Уменьшение объема или массы скелетной мышцы называется атрофией. Уменьшение объема или массы скелетной мышцы в пожилом возрасте называется саркопенией.
Гипертрофия обуславливает скорость сокращения скелетной мышцы, максимальную силу, а также способность противостоять утомлению, — все это важные физические качества, имеющие непосредственное отношение к спортивным показателям. Благодаря высокой вариативности различных характеристик мышечной ткани, таких, как размер и состав мышечных волокон, а также степень капилляризации ткани, скелетные мышцы способны быстро приспосабливаться к изменениям, возникающим в ходе тренировочного процесса. В то же время характер адаптации скелетных мышц к силовым упражнениям и упражнениям на выносливость будет отличаться, что свидетельствует о существовании различных систем реагирования на нагрузку.
Таким образом, приспособительный процесс скелетных мышц к тренировочным нагрузкам можно рассматривать как совокупность согласованных локальных и периферических событий, ключевыми регуляторными сигналами к которым являются гормональные, механические, метаболические и нервные факторы. Изменения в скорости синтеза гормонов и ростовых факторов, а также содержание их рецепторов являются важными факторами регуляции приспособительного процесса, позволяющего скелетным мышцам удовлетворить физиологические потребности различных видов двигательной активности.
Типы гипертрофии мышечных волокон [ править | править код ]
Можно выделить два крайних типа гипертрофии мышечных волокон [1] [2] : миофибриллярную гипертрофию и саркоплазматическую гипертрофию.
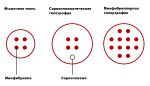
Нередко к гипертрофии относят и гиперплазию мышцы (увеличение количества волокон), однако последние исследования [4] показали, что вклад гиперплазии в объем мышцы составляет менее 5% и носит более существенный характер только при использовании анаболических стероидов. Гормон роста при этом не вызывает гиперплазии. Таким образом, люди склонные к гипертрофии, как правило, имеют большее количество мышечных волокон. Общее число волокон заложено генетически и практически не меняется в течение жизни без применения специальной фармакологии.
Методика оценки степени гипертрофии [ править | править код ]
Для того, чтобы оценить степень гипертрофии скелетной мышцы, необходимо измерить изменение её объема или массы. Современные методы исследования (компьютерная или магнито-резонансная томография) позволяют оценить изменение объема скелетных мышц человека и животных. С этой целью выполняются многократные «срезы» поперечного сечения мышцы, что позволяет вычислить её объем. Однако, до настоящего времени о степени гипертрофии скелетных мышц достаточно часто судят по изменению максимального значения поперечного сечения мышцы, полученного посредством компьютерной или магниторезонансной томографии.
В бодибилдинге гипертрофию мышц оценивают измеряя охваты рук (на уровне предплечья и бицепса), бедер, голеней, грудной клетки с помощью метровой ленты.
Показатели, определяющие объем скелетных мышц [ править | править код ]
В первом приближении [6] объем всей мышцы (Vм) можно выразить формулой:
Влияние тренировки на параметры, определяющие объем скелетных мышц [ править | править код ]
Механизмы гипертрофии скелетных мышц [ править | править код ]
В основе миофибриллярной гипертрофии мышечных волокон лежит интенсивный синтез и уменьшенный распад мышечных белков. Существует несколько гипотез миофибриллярной гипертрофии:
Гипотеза ацидоза предполагает, что пусковым стимулом для повышенного синтеза белка в скелетных мышцах является накопление в них молочной кислоты (лактата). Увеличение лактата в мышечных волокнах вызывает повреждение сарколеммы мышечных волокон и мембран органелл, появление в саркоплазме мышечных волокон ионов кальция, что вызывает активизацию протеолитических ферментов, расщепляющих мышечные белки. Увеличение синтеза белка в этой гипотезе связано с активацией и последующим делением клеток-сателлитов.
Гипотеза гипоксии предполагает, что пусковым стимулом для повышенного синтеза белка в скелетных мышцах является временное ограничение поступления кислорода (гипоксия) к скелетным мышцам, что происходит при выполнении силовых упражнений с большими отягощениями. Гипоксия и последующая реперфузия (восстановление притока кислорода к скелетным мышцам) вызывает повреждение мембран мышечных волокон и органоидов, появление в саркоплазме мышечных волокон ионов кальция, что вызывает активизацию протеолитических ферментов, расщепляющих мышечные белки. Увеличение синтеза белка в этой гипотезе связано с активизацией и последующим делением клеток-сателлитов.
Согласно вышеописанным гипотезам повреждение мышечного волокна вызывает запаздывающие болезненные ощущения в мышцах (DOMS), что связывается с их воспалением.
Возрастное развитие мышечной массы идет параллельно с увеличением продукции андрогенных гормонов. Первое заметное увеличение объема мышечных волокон наблюдается в 6-7-летнем возрасте, когда усиливается образование андрогенов. С наступлением полового созревания (11 – 15 лет) начинается интенсивный прирост мышечной массы у мальчиков, который продолжается и после периода полового созревания. У девочек развитие мышечной массы в основном заканчивается с периодом полового созревания.
В опытах на животных установлено, что введение препаратов андрогенных гормонов (анаболиков) вызывает значительную интенсификацию синтеза мышечных белков, в результате чего увеличивается масса тренируемых мышц и как результат – их сила. Вместе с тем гипертрофия скелетных мышц может происходить и без участия андрогенных и других гормонов (гормона роста, инсулина и тиреоидных гормонов).
Влияние тренировки на композицию и гипертрофию мышечных волокон различных типов.
В результате силовой тренировки степень гипертрофии быстрых мышечных волокон (II типа) значительно больше, чем медленных волокон (I типа), тогда как тренировка направленная на выносливость ведет к гипертрофии в первую очередь медленных волокон (I типа). Эти различия показывают, что степень гипертрофии мышечного волокна зависит, как от меры его использования в процессе тренировок, так и от его способности к гипертрофии.
Силовая тренировка связана с относительно небольшим числом повторных максимальных или близких к ним мышечных сокращений, в которых участвуют как быстрые, так и медленные мышечные волокна. Однако и небольшого числа повторений достаточно для развития гипертрофии быстрых волокон, что указывает на их большую предрасположенность к гипертрофии (по сравнению с медленными волокнами). Высокий процент быстрых волокон (II типа) в мышцах служит важной предпосылкой для значительного роста мышечной силы при направленной силовой тренировке. Поэтому люди с высоким процентом быстрых волокон в мышцах имеют более высокие потенциальные возможности для развития силы и мощности.
Тренировка выносливости связана с большим числом повторных мышечных сокращений относительно небольшой силы, которые в основном обеспечиваются активностью медленных мышечных волокон. Поэтому при тренировке на выносливость более выражена гипертрофия медленных мышечных волокон (I типа) по сравнению с гипертрофией быстрых волокон (II типа).
Факторы гипетрофии [ править | править код ]
Синтез сократительных белков [ править | править код ]
Рибонуклеиновая кислота (РНК) [ править | править код ]
Гиперплазия (клетки-сателлиты) [ править | править код ]
Клетки-сателлиты или спутниковые клетки
Влияние андрогенных анаболических стероидов [ править | править код ]
Влияние инсулина, аминокислот и физических упражнений на гипертрофию [ править | править код ]

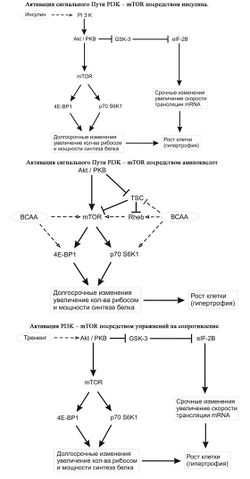
Гипертрофия скелетных мышц человека под воздействием различных средств и методов силовой тренировки
Рассмотрены механизмы гипертрофии мышц человека под влиянием отягощений различной массы, различных режимов мышечного сокращения и метода тренировки «до отказа». Предложен системный подход для описания влияния различных средств и методов тренировки на гипертрофию мышц человека. Показано, что основной механизм гипертрофии мышц связан с повреждением мышечных волокон, их последующей регенерацией и гипертрофией.
Самсонова, А.В. Гипертрофия скелетных мышц человека под воздействием различных средств и методов силовой тренировки /А.В.Самсонова // Научно-педагогические школы университета. Научные труды. Ежегодник 2014.- СПб: НГУ им. П.Ф.Лесгафта.- С.11-23.
Самсонова А.В.
Гипертрофия скелетных мышц человека под воздействием различных средств и методов силовой тренировки
Введение
Гипертрофия скелетных мышц (увеличение их массы или объема) является предметом исследования многих научных дисциплин, таких как: анатомия, гистология, биохимия, физиология, спортивная медицина, биомеханика и конечно, атлетизм. Благодаря тому накоплен большой арсенал научных знаний об этом феномене. Столь пристальное внимание к этому феномену связано с тем, что гипертрофия скелетных мышц лежит в основе увеличении их силы. Однако анализ существующей научно-методической литературы свидетельствует о том, что до настоящего времени отсутствует целостная концепция, объясняющая механизм воздействия физической нагрузки на гипертрофию скелетных мышц. В связи с этим цель исследования состояла в разработке концепции гипертрофии скелетных мышц под воздействием различных средств и методов силовой тренировки с позиций системного подхода.
Результаты
Уже в середине ХХ века в реабилитационной медицине был установлен факт быстрого восстановления массы и силы скелетных мышц после их атрофии посредством тренировки с использованием значительных внешних отягощений (70-80% от максимума). Тренировка с малыми отягощениями (30-40% от максимума), такого эффекта не вызывала (T.L. DeLorme, 1945). Найденная эмпирическим путем закономерность в настоящее время активно используется в атлетизме (А.Н. Воробьев, 1988; V.M. Zatsiorsky, W.J. Kraemer, 2006; Г.П. Виноградов, 2009). Чтобы понять механизм воздействия отягощений различной массы на увеличение объема скелетных мышц необходимо привлечь знания из ряда медико-биологических дисциплин. Из анатомии и гистологии известно, что скелетные мышцы человека состоят из мышечных волокон (МВ) различного типа: медленных (I типа) и быстрых (IIA и IIВ типа). МВ I типа характеризуются невысокой скоростью и силой сокращения, однако способность сопротивляться утомлению у них очень высока. Мышечные волокна IIВ типа способны сокращаться с большой силой и скоростью, однако сопротивление утомлению у них низкое. Волокна IIA типа характеризуются промежуточными свойствами. Основу сократительного аппарата мышечных волокон скелетных мышц человека составляют миофибриллы, которые состоят из секций – саркомеров, отделенных друг от друга Z-дисками. Гистологами установлено, что медленные волокна I типа имеют более толстые Z-диски по сравнению с быстрыми волокнами II типа (H. Hoppeler, 1986). Между собой миофибриллы на уровне Z-дисков связаны цитоскелетными белками. Помимо этого периферические миофибриллы прикреплены к внутренней оболочке мышечного волокна посредством костамеров (рис.1). Саркомер подобен связке шестигранных карандашей (в саркомере их больше 300), грифелем которых служит толстый филамент, а ребрами – тонкие филаменты. При сокращении мышцы тонкие филаменты скользят относительно толстых, расстояние между Z-дисками уменьшается, саркомер укорачивается. Одновременное сокращение всех саркомеров приводит к уменьшению длины миофибриллы и мышечного волокна. Ввиду того, что саркомер представляет собой не плоскую, а объемную структуру, при его сокращении происходит также увеличение площади его поперечного сечения (когда тонкие нити входят в промежутки между толстыми), площади поперечного сечения мышечных волокон и всей мышцы. Физиологами найдено, что управление мышцей со стороны ЦНС осуществляется посредством активации двигательных единиц (ДЕ). По классификации Р. Берка с соавт. (R.E. Burke et all. 1973) ДЕ делятся на три типа: S (slow) – медленные, устойчивые к утомлению; FR (fast resistant) – быстрые, устойчивые к утомлению, FF – fast fatigable – быстрые, быстроутомляемые. ДЕ различных типов соответствуют различные виды мышечных волокон. В состав двигательных единиц типа S входят мышечные волокна I типа. В состав двигательных единиц типа FR – IIA типа. В состав двигательных единиц типа FF – входят мышечные волокна IIB типа.
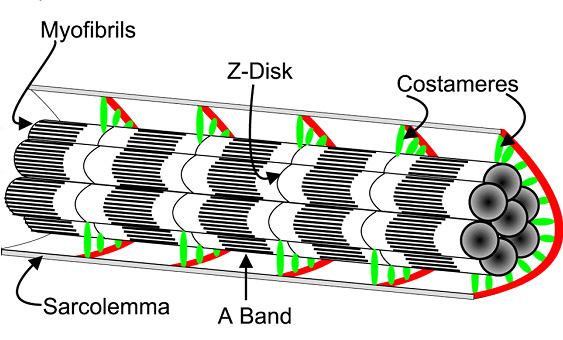 Рис.1. Прикрепление миофибрилл посредством костамеров к сарколемме мышечного волокна (J.M.Ervasty, 2003)
Рис.1. Прикрепление миофибрилл посредством костамеров к сарколемме мышечного волокна (J.M.Ervasty, 2003)
Количество ДЕ, активных в процессе сокращения мышцы, определяется посредством центральных и рефлекторных механизмов регуляции силы мышц (А.Г. Фельдман, 1979). Установлено, что имеется стабильный порядок вовлечения в работу (рекрутирования) ДЕ: вначале рекрутируются ДЕ S типа, затем FR типа, последними в сокращение вовлекаются ДЕ FF типа (E. Henneman, C.B. Olson, 1965). Концепция, объясняющая воздействие больших отягощений на увеличение силы и гипертрофию скелетных мышц состоит в следующем. Если внешнее отягощение небольшое (например, масса штанги составляет менее 20% максимума) при выполнении силового упражнения рекрутируются только мышечные волокна I типа, при этом уровень силы, развиваемый мышцей, невысокий (В.С. Гурфинкель, Ю.С. Левик, 1985). Для преодоления большого внешнего отягощения (более 70% от максимума) мышца должна развить большую силу. Поэтому в сокращение последовательно вовлекаются все типы мышечных волокон, в том числе и IIB типа (рис.2).
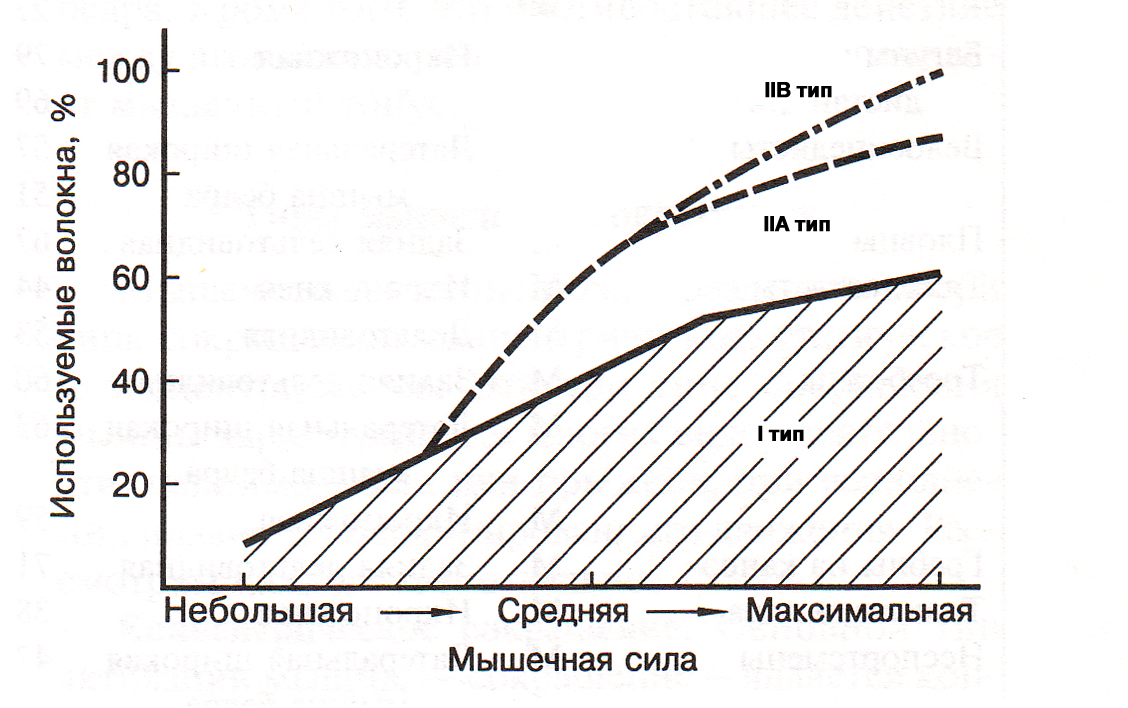 Рис.2. Зависимость между развиваемой силой мышцы и вовлечением в работу медленных и быстрых
Рис.2. Зависимость между развиваемой силой мышцы и вовлечением в работу медленных и быстрых
мышечных волокон (Дж. Х.Уилмор, Д.Л. Костилл, 1997)
Специалистами в области спортивной медицины установлено, что тренировка с применением больших отягощений вызывает мышечные боли у спортсменов, как во время, так и после ее окончания (Б.И. Прилуцкий, 1989; В.И. Морозов, Г.А. Сакута, М.И. Калинский, 2006; Г.А. Макарова, 2008). В настоящее время накоплено достаточное количество фактов, свидетельствующих о том, что болезненные ощущения в мышцах в первую очередь связаны с их повреждением. Повреждение мышечных волокон вызывает воспаление, что ощущается как боль в мышцах через 24 часа и более. Последующая регенерация мышечных волокон приводит к их гипертрофии.
Различают несколько режимов сокращения мышц: преодолевающий (концентрический) – длина мышцы уменьшается, уступающий (эксцентрический) – длина мышцы увеличивается, изометрический (статический) – длина мышцы не изменяется. P.J. Rasch и L.J. Morehouse (1957) одними из первых показали, что динамические упражнения по сравнению с изометрическими вызывают больший прирост силы и площади поперечного сечения мышц. Показано, что тренировка с использованием эксцентрического режима приводит к несколько большей гипертрофии мышц, чем другие режимы сокращения (M.J.Gibala et al. 1995; Е. Hagbie et al. 1996; J.Y. Seger, B. Arvidsson, A. Thorstensson, 1998; M.J. Gibala et al. 2000). Установлено, что гипертрофия мышечных волокон при работе в эксцентрическом режиме больше, чем при концентрическом (S.L.Lindstedt, P.C. LaStayo, T.E. Reich, 2001).
Исследования, проведенные на гистологическом уровне, свидетельствуют о том, что при выполнении упражнений в эксцентрическом режиме в большей степени повреждается цитоскелет и Z-диски мышечного волокна по сравнению с тренировкой в других режимах (J. Friden, M. Sjostrom, B. Ekblom, 1983; J.Friden, R.L. Lieber, 2001), рис.3. Установлено, что степень повреждения Z-дисков мышечных волокон II типа в три раза больше, чем у волокон I типа (J. Friden, M. Sjostrom, B. Ekblom, 1983).
Рис.3. Электронная фотография мышечного волокна человека после выполнения эксцентрических упражнений. Обозначения: * — миофибриллы с разрушенными Z-дисками; ○ – миофибрилла с неповрежденным Z диском. (J.G. Yu, L.Carlsson, L.E. Thomell, 2004)
Исследованиями M.J. Gibala etal. (1995) показано, что даже однократная силовая тренировка в эксцентрическом режиме вызывает у начинающих спортсменов повреждение более 82% мышечных волокон, а в концентрическом – только 33%, при этом во время эксцентрических сокращений суммарная ЭАМ на 40% меньше, чем во время концентрических. У хорошо тренированных спортсменов аналогичная тренировка приводит к 45% повреждений мышечных волокон при работе в эксцентрическом режиме и 27% при работе в концентрическом режиме (M.J. Gibala et al. 2000). Биохимические данные свидетельствуют о том, что на третий-пятый день после эксцентрических сокращений, в крови исследуемых значительно возрастает уровень креатинкиназы и миоглобина, что свидетельствует о сильных повреждениях мышечных волокон. При этом он значительно превышает аналогичные показатели, полученные при выполнении движений в концентрическом режиме (J.A. Faulkner, S.V. Brooks, J.A. Opiteck, 1993; A.P. Lavender, K. Nosaka, 2006; K. Nosaka, 2008). На основе изучения биомеханических характеристик развития усилия мышцей M.J. Gibala et al. (1995) было показано (рис.4), что после одного тренировочного занятия силовыми упражнениями значения максимального момента силы двуглавой мышцы плеча, измеренного в изометрическом режиме, понизились как у руки, выполнявшей движения в концентрическом, так и у руки, выполнявшей движения в эксцентрическом режимах сокращения (р≤0,05). Однако через 24 часа этот показатель, измеренный у руки, работающей в концентрическом режиме достоверно не отличался от базового уровня (р>0,05). В то же время значения максимального момента силы, у руки, выполняющей эксцентрические сокращения, достоверно отличались от начального уровня через 24, 48, 72 и даже 96 часов (р≤0,05). Следует отметить, что растяжение пассивной мышцы не приводит к ее гипертрофии (J.R. Fowles et al. 2000).
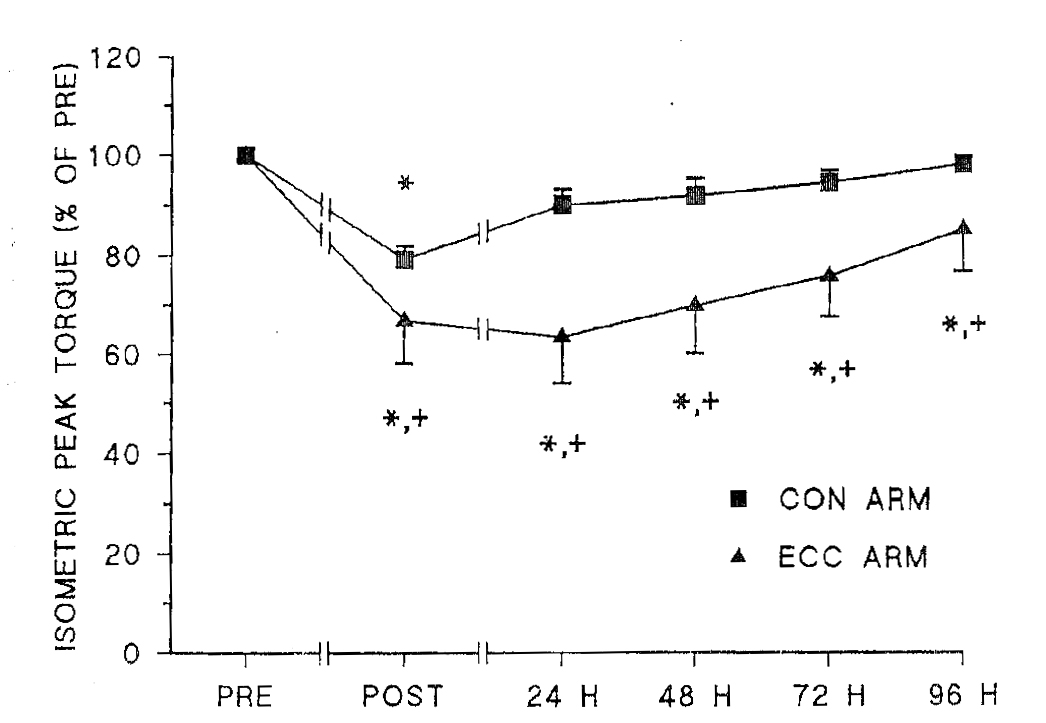 Рис. 4. Максимальные значения момента силы, полученного в изометрическом режиме для руки, выполнявшей концентрические сокращения (квадраты) и руки, выполнявшей эксцентрические сокращения (треугольники) в различные моменты времени: до (PRE), сразу после (POST),через 24 часа (24H),48 часов (48H), 72 часа (72H) и 96 часов (96H) после выполнения упражнения. Представлены средние значения и ошибка среднего до и после выполнения упражнения * – различия между значениями, полученными до и после проведения упражнений достоверны на уровне значимости р≤0,05. + – различия статистически достоверны на уровне значимости р≤0,05 между CON рукой и Eсс рукой (по: M.J.Gibala et al. (1995).
Рис. 4. Максимальные значения момента силы, полученного в изометрическом режиме для руки, выполнявшей концентрические сокращения (квадраты) и руки, выполнявшей эксцентрические сокращения (треугольники) в различные моменты времени: до (PRE), сразу после (POST),через 24 часа (24H),48 часов (48H), 72 часа (72H) и 96 часов (96H) после выполнения упражнения. Представлены средние значения и ошибка среднего до и после выполнения упражнения * – различия между значениями, полученными до и после проведения упражнений достоверны на уровне значимости р≤0,05. + – различия статистически достоверны на уровне значимости р≤0,05 между CON рукой и Eсс рукой (по: M.J.Gibala et al. (1995).
Концепция, описывающая последовательность событий, приводящих к большему повреждению мышечных волокон при работе в эксцентрическом режиме по сравнению с концентрическим и изометрическим состоит в следующем. Для того, чтобы мышца, выполняющая статическую работу, начала удлиняться, момент внешних сил должен превосходить момент силы тяги мышцы. Это возможно только в том случае, если часть двигательных единиц будет деактивирована, то есть прекратит свою активность и, как следствие – будет уменьшено количество активных мышечных волокон. V. Eloranta, P.W. Komi (1980) и M.J.Gibala et al. (1995) находят, что этот механизм управления ДЕ лежит в основе эксцентрических сокращений мышцы. Это предположение подтверждается исследованиями метаболических затрат и суммарной ЭАМ, которые меньше при работе в эксцентрическом
режиме по сравнению с другими режимами. Вследствие деактивации части ДЕ момент внешней силы становится больше момента силы, развиваемого мышцей и, как следствие – активная мышца начинает удлиняться. Удлинение мышцы сопровождается удлинением мышечных волокон и миофибрилл, так как миофибриллы внутри мышечного волокна имеют «жесткую» привязку к его мембране посредством костамеров и элементов цитоскелета. Удлинение миофибрилл возможно только за счет увеличения длины элементов миофибрилл – саркомеров. Удлинению саркомера, находящегося в активном состоянии, препятствуют силы, возникающие между его толстыми и тонкими филаментами, которые стремятся уменьшить его длину. В связи с тем, что значение внешнего момента силы превосходит момент силы, развиваемый сократительными элементами мышцы, саркомер растягивается. Следствием этого является повреждение элементов цитоскелета и мембранного скелета мышечных волокон, а также повреждение Z-дисков миофибрилл (J. Friden, U. Kjorell, R.L. Lieber, 1984). Таким образом, большее воздействие эксцентрического режима работы на гипертрофию скелетных мышц связано также с их повреждением.
3. Влияние на гипертрофию скелетных мышц силовой тренировки методом «до отказа»
Метод повторных непредельных усилий (до «отказа») – один из методов увеличения силовых способностей спортсменов (Ю.Ф. Курамшин, 2004). Используя гистологические методы В.Ф. Кондаленко (1976) обнаружил серьезные повреждения миофибрилл после однократной физической нагрузки, выполненной до «отказа». Если однократная нагрузка проводилась до глубокого утомления, то деструктивные изменения в миофибриллах сохранялись и через 96 часов после ее окончания. Физиологическое исследование (Е.Б. Мякинченко, В.Н. Селуянов, 2005) работы мышц до «отказа» при выполнении медленных приседаний со штангой 50-60% от максимума показало, что амплитуда суммарной ЭМГ на протяжении всего подхода была ниже максимальной. В момент «отказного» повторения она увеличивалась. Факты серьезного повреждения мышечных волокон подтверждаются данными А.Д. Минигалина с соавт. (2011), которые изучали срочные и отставленные биохимические и физиологические эффекты прямой мышцы бедра и организма в целом после однократной физической нагрузки на силовом тренажере методом до «отказа». Ими установлена значительная активность фермента креатинкиназы через трое суток после выполнения тренировочного задания. Авторы находят, что такая динамика активности фермента свидетельствует о значительной его утечке из мышечных волокон, что говорит об их повреждениях. Кроме того, все исследуемые испытывали сильные болезненные ощущения в мышцах, максимум которых приходился на 2-3 сутки после физической нагрузки. Нами было проведено исследование ряда биомеханических и физиологических показателей четырехглавой мышцы бедра и ее головки (латеральной широкой мышцы бедра) при выполнении силовых упражнений методом до «отказа», в котором участвовали спортсмены различной квалификации (Самсонова А.В., 2010, 2011; Самсонова А.В., Косьмина Е.А., 2011). Полученные результаты позволили сделать следующие выводы:
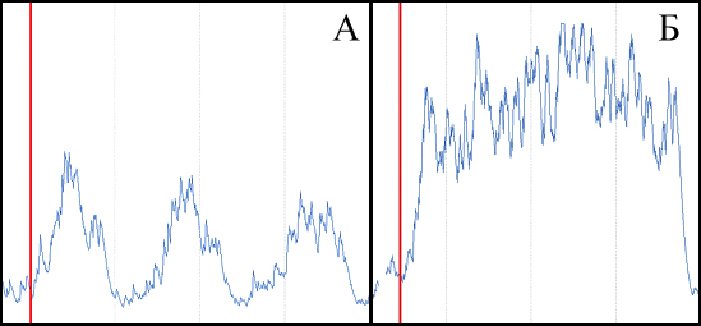 Рис. 5. Суммарная электрическая активность латеральной широкой м. бедра при выполнении 2, 3 и 4 повторения (А) и отказного повторения (Б) силового упражнения с отягощением в 40% от максимума в пятом подходе. Жирные вертикальные линии соответствуют началу второго (А) и отказного (Б) повторений упражнения (А.В. Самсонова, Е.А. Косьмина, 2011)
Рис. 5. Суммарная электрическая активность латеральной широкой м. бедра при выполнении 2, 3 и 4 повторения (А) и отказного повторения (Б) силового упражнения с отягощением в 40% от максимума в пятом подходе. Жирные вертикальные линии соответствуют началу второго (А) и отказного (Б) повторений упражнения (А.В. Самсонова, Е.А. Косьмина, 2011)
Следует отметить, что долговременные адаптационные сдвиги в организме при использовании метода до «отказа» зависят от значений внешней нагрузки и квалификации исследуемых. Предлагаемая нами концепция воздействия физической нагрузки методом до «отказа» с внешним отягощением 70-80% от максимума состоит в следующем. Рассмотрим для примера выполнение силового упражнения «жим штанги ногами лежа на тренажере» с отягощением, составляющим 80% от максимума. Физическая нагрузка состоит в выполнении 5 подходов методом «до отказа». В первом подходе на выполнение восьми повторений спортсмен затрачивает в среднем 20-30 с. К концу первого подхода в «отказном» повторении количество мышечных волокон, способных развивать необходимое усилие, резко уменьшается – из-за того, что к 30-45 с запасы креатинфосфата в мышце подходят к концу (Н.И. Волков, 2000). Поэтому ЦНС в последних циклах движения «бросает в бой» свой резерв – самые большие и сильные ДЕ, которые еще не принимали участие в работе и потому сохранили достаточное количество этого энергетического вещества. Этим можно объяснить увеличение амплитуды ЭМГ в «отказном» повторении первого подхода. Однако падение уровня силы, которую способна развить мышца при «отказном» повторении первого подхода из-за истощения энергетических ресурсов мышечных волокон очень большое. Поэтому мышца уже не способна развить требуемый импульс силы (произведение силы мышцы на длительность ее активности), чтобы сообщить внешнему отягощению необходимую скорость после его остановки в нижнем положении. В связи с этим, выполнение упражнения в первом подходе прекращается. Через несколько минут отдыха в части истощенных мышечных волокон запасы креатинфосфата частично восстанавливаются и мышца снова может развить достаточное усилие. Поэтому спортсмен способен выполнить следующий подход до «отказа». Но так как часть мышечных волокон уже истощена или повреждена, количество повторений в подходе «до отказа» уменьшается. Если спортсмен выполняет не более пяти подходов методом до «отказа», повреждение мышечных волокон невелико, о чем свидетельствует восстановление уровня максимальной изометрической силы через 24 часа после нагрузки. Если спортсмен выполняет шесть и более подходов, уровень силы, развиваемый мышцей, резко падает, развитие необходимого импульса силы происходит за счет значительного увеличения длительности «отказного» подхода. При этом скорость сокращения мышцы в концентрическом режиме резко уменьшается, и мышца начинает функционировать практически в изометрическом режиме, который, по сравнению с концентрическим, обладает большим повреждающим воздействием на мышечные волокна. Поэтому при медленном выполнении последнего, «отказного» повторения в последнем подходе происходит значительное повреждение большого количества мышечных волокон. К концу последнего подхода в «отказном» повторении оставшиеся активными ДЕ уже не способны развить необходимый импульс силы, чтобы сообщить внешнему отягощению (80% от максимума) необходимую скорость. Спортсмен прекращает выполнение упражнения. Если работа до отказа выполняется с небольшим внешним отягощением, например, 40% от максимума, то к последнему, «отказному» повторению последнего подхода большая часть мышечных волокон отключается не из-за повреждения, а из-за истощения вследствие того, что в них исчерпались запасы энергии. Работа в таком режиме в большей степени ведет к развитию силовой выносливости, чем силы. Таким образом, изложенная концепция позволяет объяснить большинство наблюдаемых эффектов в работе до «отказа», важным моментом которой является активация и повреждение больших ДЕ, в состав которых входят МВ II типа.
Выводы и заключение
Предложен системный подход для описания влияния различных средств и методов тренировки на гипертрофию скелетных мышц человека. Показано, что основной механизм гипертрофии скелетных мышц связан с повреждением мышечных волокон и их последующей регенерацией и гипертрофией. Во время выполнения силовых упражнений не все мышечные волокна активны. Если мышечное волокно не активно, оно не повреждается. Для того, чтобы активизировать мышечные волокна II типа, которые дают максимальный прирост площади поперечного сечения мышцы при выполнении силовых упражнений необходимо использовать или большие отягощения или метод «до отказа». Также сильно мышечные волокна II типа повреждаются при выполнении силовых упражнений в эксцентрическом (уступающем режиме). Гипотеза повреждения мышечных волокон и их последующей регенерации подтверждается фактами, полученными представителями спортивной медицины о наличии болезненных ощущений в мышцах при выполнении силовых упражнений с большими отягощениями во время и после окончания тренировочного занятия (синдром DOMS).



